

Intracellular Calcium Mobilization Is Required for Sonic Hedgehog Signaling
- PMID: 29754802
- PMCID: PMC6007892
- DOI: 10.1016/j.devcel.2018.04.013
Intracellular Calcium Mobilization Is Required for Sonic Hedgehog Signaling
Abstract
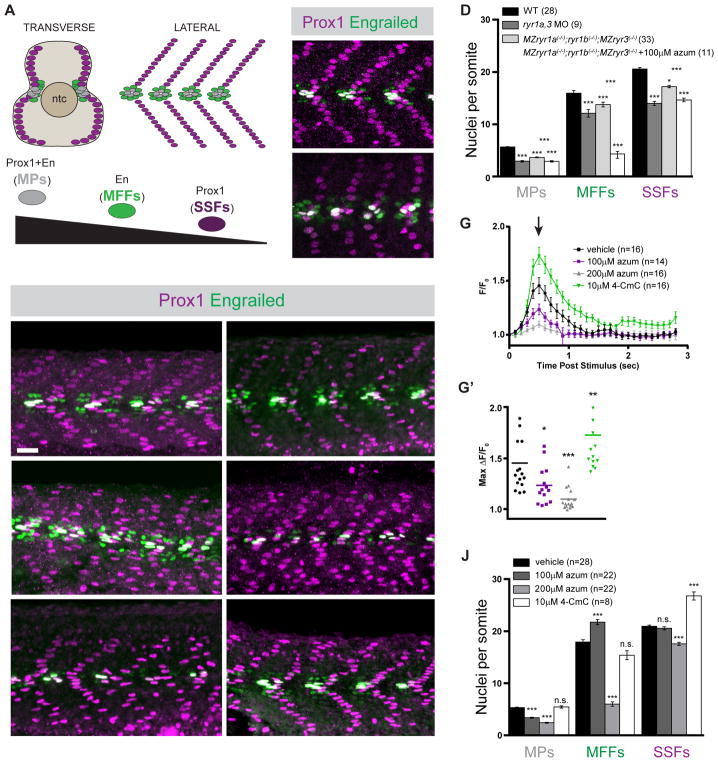
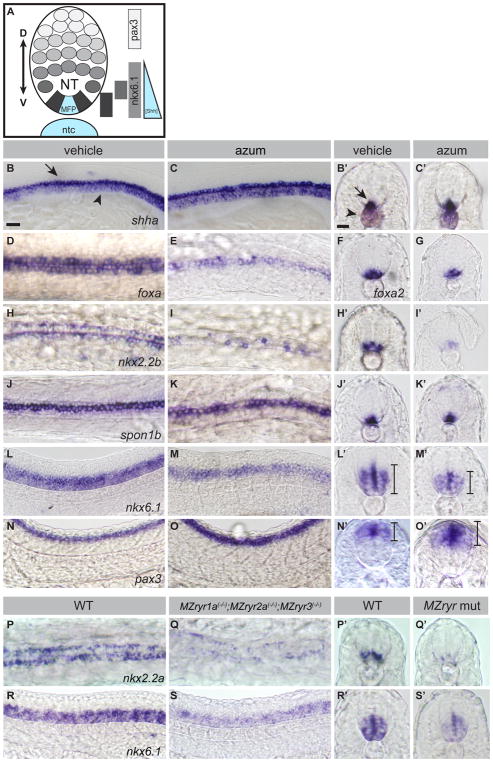
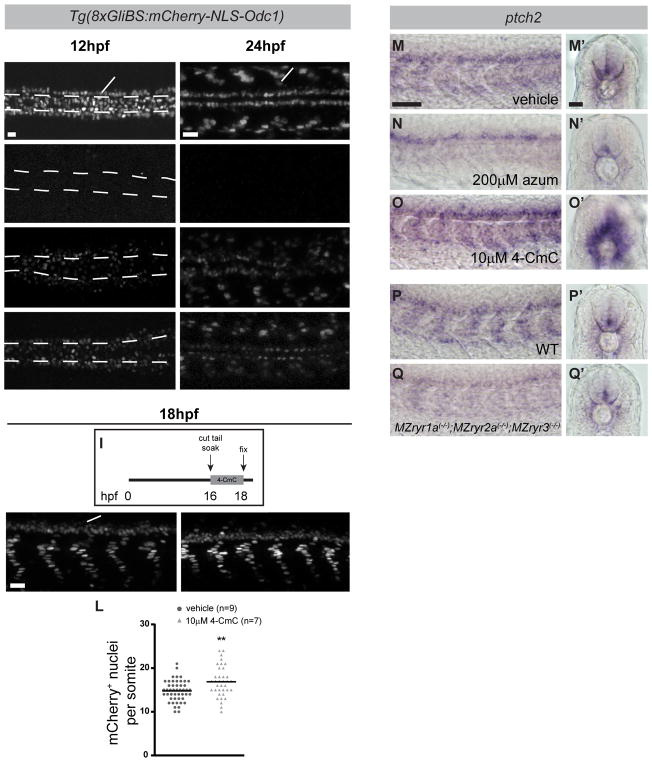
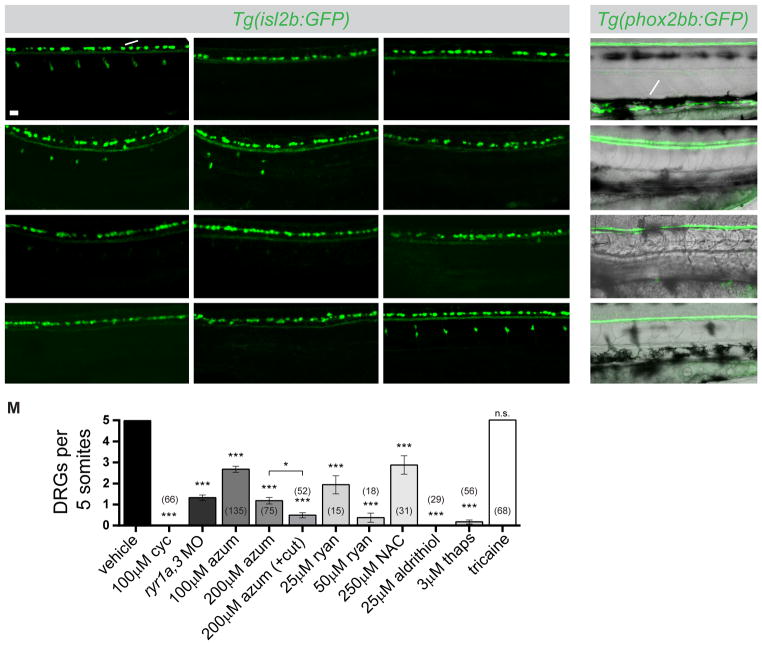
Graded Shh signaling across fields of precursor cells coordinates patterns of gene expression, differentiation, and morphogenetic behavior as precursors form complex structures, such as the nervous system, the limbs, and craniofacial skeleton. Here we discover that intracellular calcium mobilization, a process tightly controlled and readily modulated, regulates the level of Shh-dependent gene expression in responding cells and affects the development of all Shh-dependent cell types in the zebrafish embryo. Reduced expression or modified activity of ryanodine receptor (RyR) intracellular calcium release channels shifted the allocation of Shh-dependent cell fates in the somitic muscle and neural tube. Mosaic analysis revealed that RyR-mediated calcium mobilization is required specifically in Shh ligand-receiving cells. This work reveals that RyR channels participate in intercellular signal transduction events. As modulation of RyR activity modifies tissue patterning, we hypothesize that alterations in intracellular calcium mobilization contribute to both birth defects and evolutionary modifications of morphology.
Keywords: hedgehog signaling; intracellular calcium release; ryanodine receptors; tissue pattering.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
The ADAMTS5 Metzincin Regulates Zebrafish Somite Differentiation.Int J Mol Sci. 2018 Mar 7;19(3):766. doi: 10.3390/ijms19030766. Int J Mol Sci. 2018. PMID: 29518972 Free PMC article.
-
Mediolateral patterning of somites: multiple axial signals, including Sonic hedgehog, regulate Nkx-3.1 expression.Mech Dev. 1998 Jan;70(1-2):25-34. doi: 10.1016/s0925-4773(97)00168-8. Mech Dev. 1998. PMID: 9510022
-
Hedgehog signaling is required for differentiation of endocardial progenitors in zebrafish.Dev Biol. 2012 Jan 15;361(2):377-91. doi: 10.1016/j.ydbio.2011.11.004. Epub 2011 Nov 12. Dev Biol. 2012. PMID: 22119054
-
Establishing and interpreting graded Sonic Hedgehog signaling during vertebrate neural tube patterning: the role of negative feedback.Cold Spring Harb Perspect Biol. 2009 Aug;1(2):a002014. doi: 10.1101/cshperspect.a002014. Cold Spring Harb Perspect Biol. 2009. PMID: 20066087 Free PMC article. Review.
-
Primary cilia and graded Sonic Hedgehog signaling.Wiley Interdiscip Rev Dev Biol. 2012 Sep-Oct;1(5):753-72. doi: 10.1002/wdev.43. Epub 2012 Apr 4. Wiley Interdiscip Rev Dev Biol. 2012. PMID: 23799571 Review.
Cited by
-
Highly Efficient CRISPR-Cas9-Based Methods for Generating Deletion Mutations and F0 Embryos that Lack Gene Function in Zebrafish.Dev Cell. 2019 Dec 2;51(5):645-657.e4. doi: 10.1016/j.devcel.2019.10.004. Epub 2019 Nov 7. Dev Cell. 2019. PMID: 31708433 Free PMC article.
-
Mutations in phospholipase C eta-1 (PLCH1) are associated with holoprosencephaly.J Med Genet. 2022 Apr;59(4):358-365. doi: 10.1136/jmedgenet-2020-107237. Epub 2021 Apr 5. J Med Genet. 2022. PMID: 33820834 Free PMC article.
-
Cholesterylation of Smoothened is a calcium-accelerated autoreaction involving an intramolecular ester intermediate.Cell Res. 2022 Mar;32(3):288-301. doi: 10.1038/s41422-022-00622-0. Epub 2022 Feb 4. Cell Res. 2022. PMID: 35121857 Free PMC article.
-
Ryanodine receptor-active non-dioxin-like polychlorinated biphenyls cause neurobehavioral deficits in larval zebrafish.Front Toxicol. 2022 Oct 6;4:947795. doi: 10.3389/ftox.2022.947795. eCollection 2022. Front Toxicol. 2022. PMID: 36278027 Free PMC article.
-
The presynaptic glycine transporter GlyT2 is regulated by the Hedgehog pathway in vitro and in vivo.Commun Biol. 2021 Oct 18;4(1):1197. doi: 10.1038/s42003-021-02718-6. Commun Biol. 2021. PMID: 34663888 Free PMC article.
References
-
- Abzhanov A, Kuo WP, Hartmann C, Grant BR, Grant PR, Tabin CJ. The calmodulin pathway and evolution of elongated beak morphology in Darwin’s finches. Nature. 2006;442:563–567. - PubMed
-
- Barresi MJ, Stickney HL, Devoto SH. The zebrafish slow-muscle-omitted gene product is required for Hedgehog signal transduction and the development of slow muscle identity. Development. 2000;127:2189–2199. - PubMed
-
- Beachy PA, Karhadkar SS, Berman DM. Tissue repair and stem cell renewal in carcinogenesis. Nature. 2004;432:324–331. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
