

Blocking Neuronal Signaling to Immune Cells Treats Streptococcal Invasive Infection
- PMID: 29754819
- PMCID: PMC5959783
- DOI: 10.1016/j.cell.2018.04.006
Blocking Neuronal Signaling to Immune Cells Treats Streptococcal Invasive Infection
Abstract
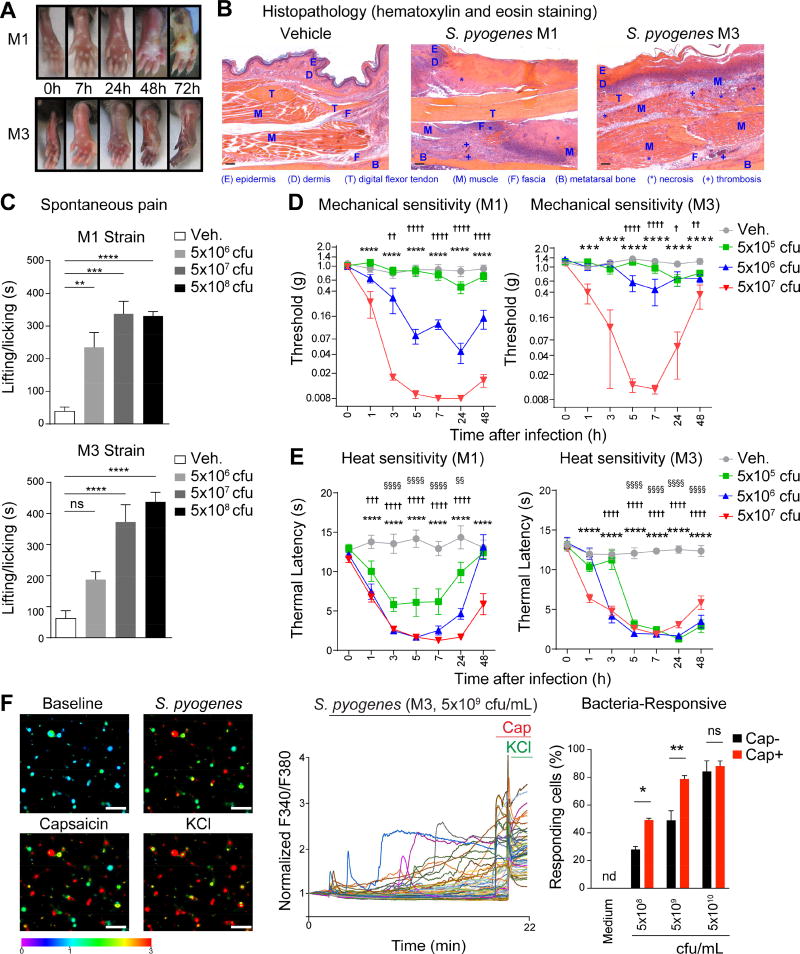
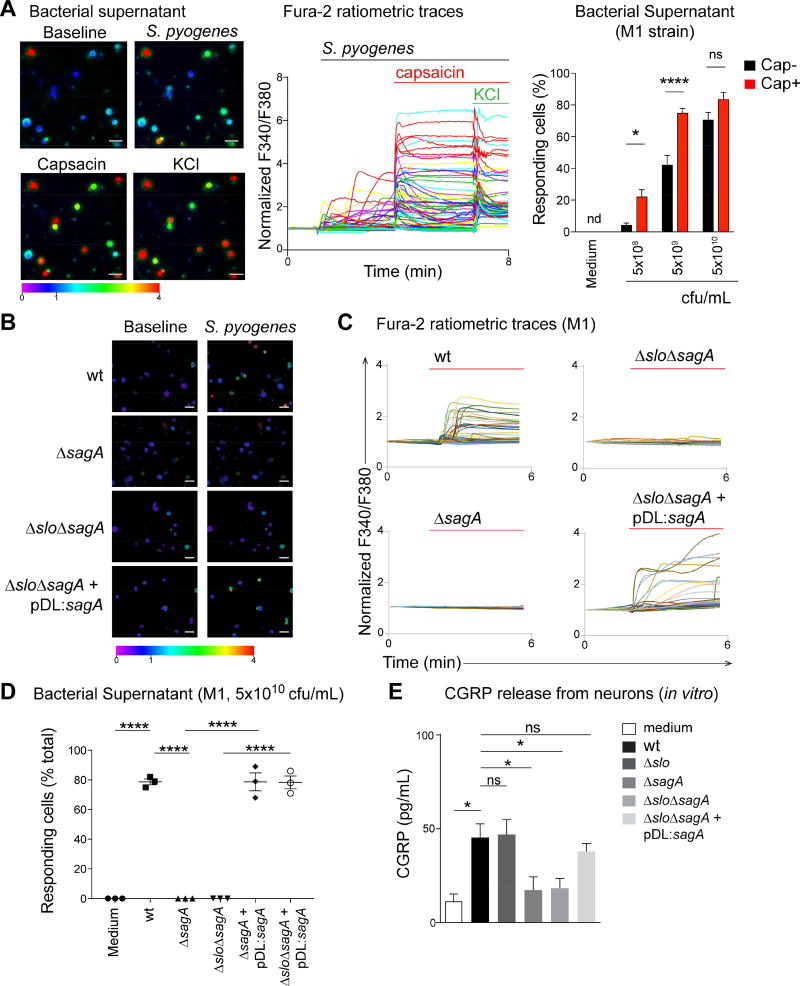
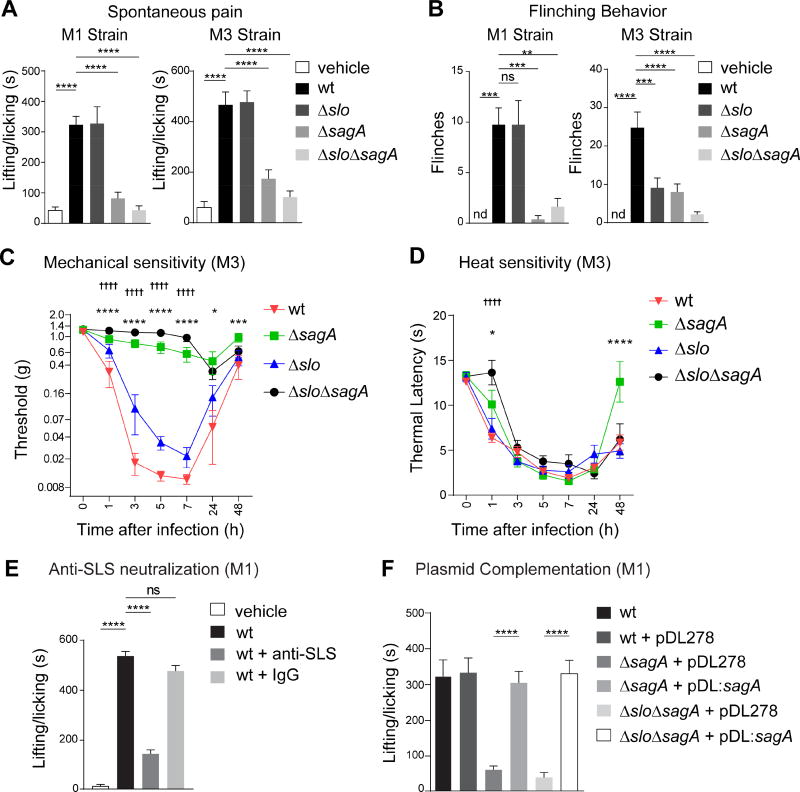
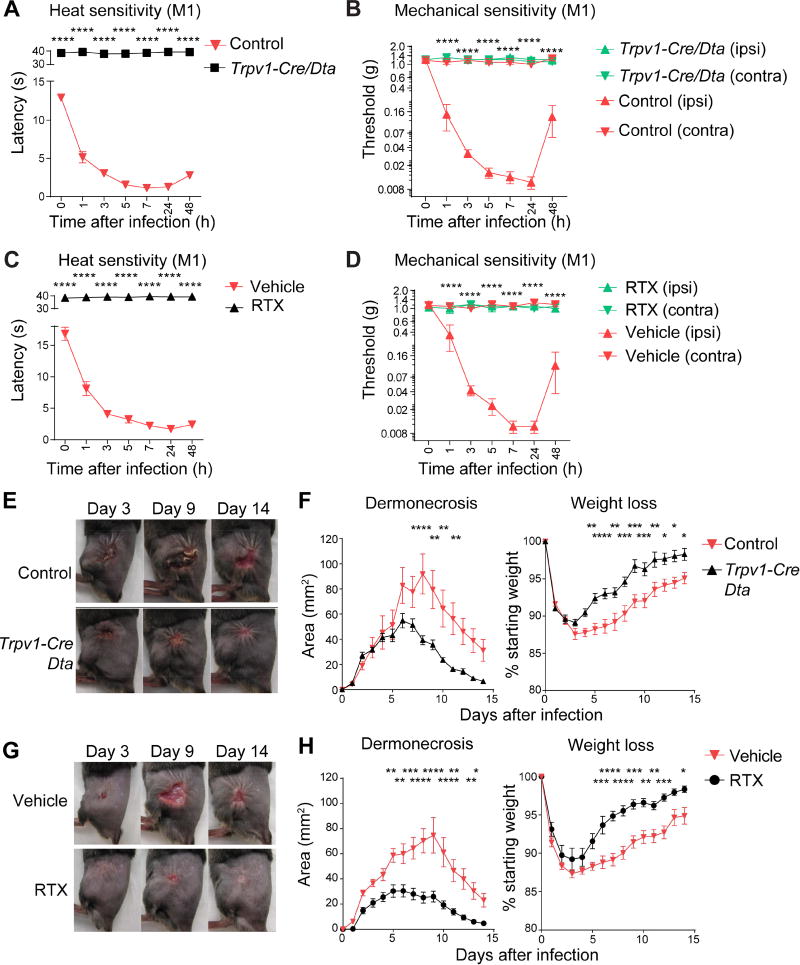
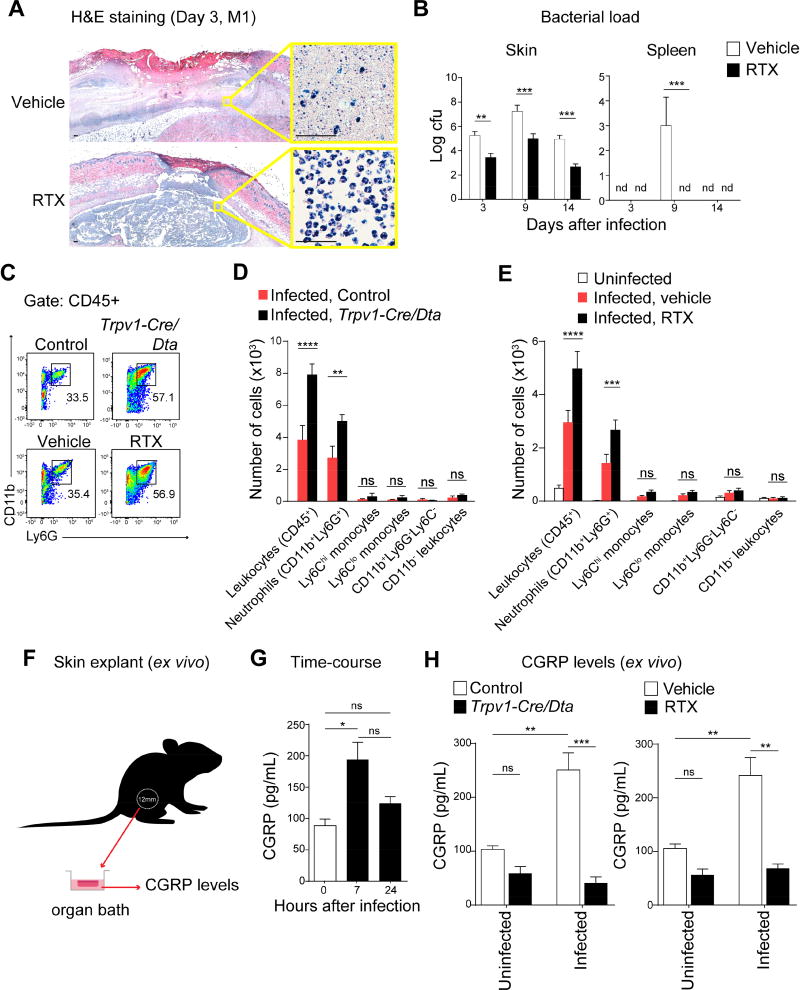
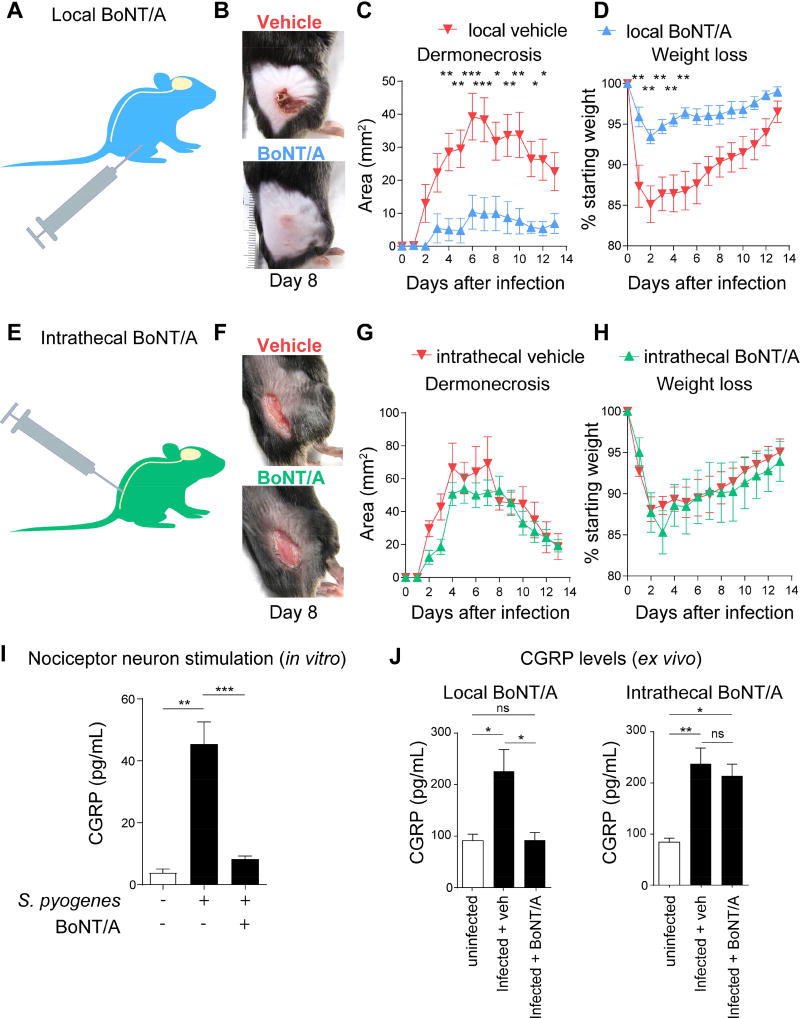
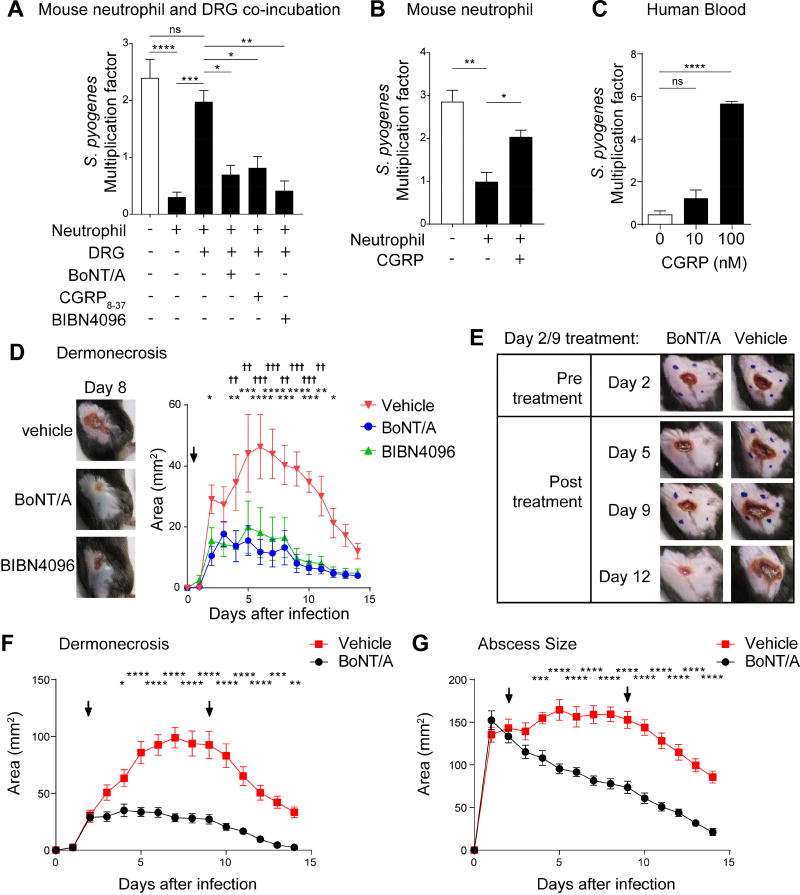
The nervous system, the immune system, and microbial pathogens interact closely at barrier tissues. Here, we find that a bacterial pathogen, Streptococcus pyogenes, hijacks pain and neuronal regulation of the immune response to promote bacterial survival. Necrotizing fasciitis is a life-threatening soft tissue infection in which "pain is out of proportion" to early physical manifestations. We find that S. pyogenes, the leading cause of necrotizing fasciitis, secretes streptolysin S (SLS) to directly activate nociceptor neurons and produce pain during infection. Nociceptors, in turn, release the neuropeptide calcitonin gene-related peptide (CGRP) into infected tissues, which inhibits the recruitment of neutrophils and opsonophagocytic killing of S. pyogenes. Botulinum neurotoxin A and CGRP antagonism block neuron-mediated suppression of host defense, thereby preventing and treating S. pyogenes necrotizing infection. We conclude that targeting the peripheral nervous system and blocking neuro-immune communication is a promising strategy to treat highly invasive bacterial infections. VIDEO ABSTRACT.
Keywords: CGRP; botulinum neurotoxin; infection; neuroimmune; neuroimmunology; neutrophil; nociceptor; pain; streptococcus pyogenes; streptolysin.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
FAPR, BB, NJY, MRW and IMC are co-inventors on patents that incorporate discoveries described in the manuscript.
Figures
Comment in
-
Neurons Are the Inflammatory Problem.Cell. 2018 May 17;173(5):1066-1068. doi: 10.1016/j.cell.2018.05.005. Cell. 2018. PMID: 29775588
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
