

Evolution of Antibiotic Resistance in Biofilm and Planktonic Pseudomonas aeruginosa Populations Exposed to Subinhibitory Levels of Ciprofloxacin
- PMID: 29760140
- PMCID: PMC6105853
- DOI: 10.1128/AAC.00320-18
Evolution of Antibiotic Resistance in Biofilm and Planktonic Pseudomonas aeruginosa Populations Exposed to Subinhibitory Levels of Ciprofloxacin
Abstract
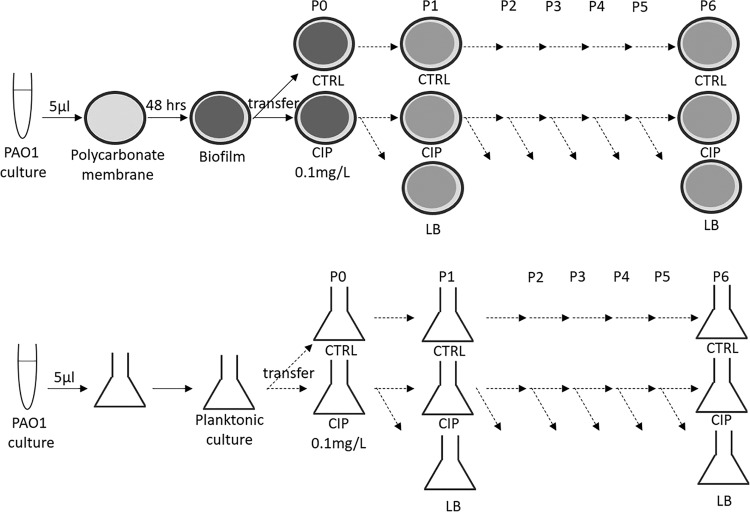
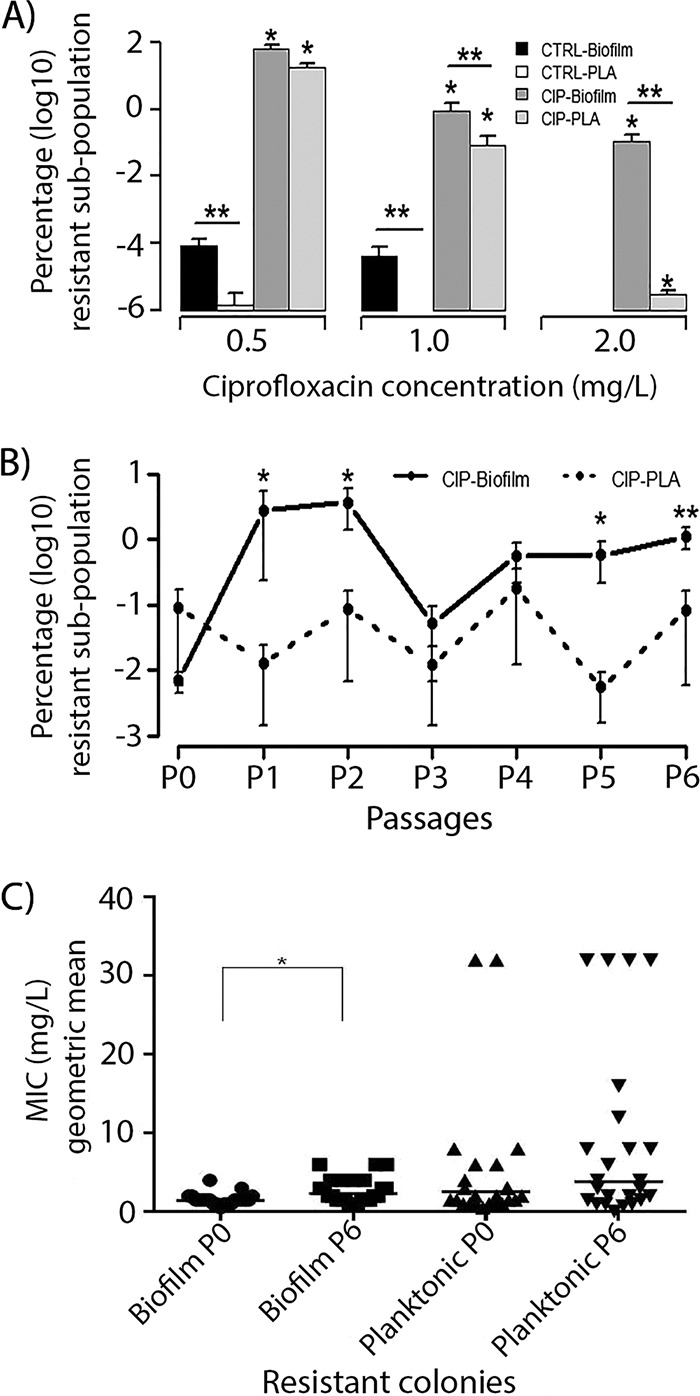
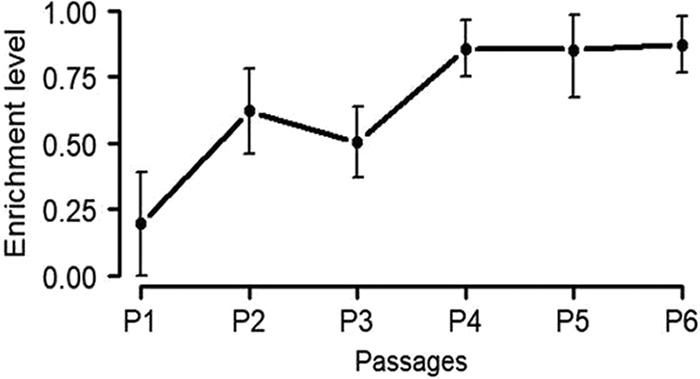
The opportunistic Gram-negative pathogen Pseudomonas aeruginosa, known for its intrinsic and acquired antibiotic resistance, has a notorious ability to form biofilms, which often facilitate chronic infections. The evolutionary paths to antibiotic resistance have mainly been investigated in planktonic cultures and are less studied in biofilms. We experimentally evolved P. aeruginosa PAO1 colony biofilms and stationary-phase planktonic cultures for seven passages in the presence of subinhibitory levels (0.1 mg/liter) of ciprofloxacin (CIP) and performed a genotypic (whole-bacterial population sequencing) and phenotypic assessment of the populations. We observed a higher proportion of CIP resistance in the CIP-evolved biofilm populations than in planktonic populations exposed to the same drug concentrations. However, the MICs of ciprofloxacin were lower in CIP-resistant isolates selected from the biofilm population than the MICs of CIP-resistant isolates from the planktonic cultures. We found common evolutionary trajectories between the different lineages, with mutations in known CIP resistance determinants as well as growth condition-dependent adaptations. We observed a general trend toward a reduction in type IV-pilus-dependent motility (twitching) in CIP-evolved populations and a loss of virulence-associated traits in the populations evolved in the absence of antibiotic. In conclusion, our data indicate that biofilms facilitate the development of low-level mutational resistance, probably due to the lower effective drug exposure than in planktonic cultures. These results provide a framework for the selection process of resistant variants and the evolutionary mechanisms involved under the two different growth conditions.
Keywords: Pseudomonas aeruginosa; biofilm; drug resistance evolution.
Copyright © 2018 American Society for Microbiology.
Figures

References
-
- Hoiby N, Bjarnsholt T, Moser C, Bassi GL, Coenye T, Donelli G, Hall-Stoodley L, Hola V, Imbert C, Kirketerp-Moller K, Lebeaux D, Oliver A, Ullmann AJ, Williams C, ESCMID Study Group for Biofilms and Consulting External Expert Werner Zimmerli . 2015. ESCMID guideline for the diagnosis and treatment of biofilm infections 2014. Clin Microbiol Infect 21(Suppl 1):S1–S25. doi: 10.1016/j.cmi.2014.10.024. - DOI - PubMed
-
- Walters MC III, Roe F, Bugnicourt A, Franklin MJ, Stewart PS. 2003. Contributions of antibiotic penetration, oxygen limitation, and low metabolic activity to tolerance of Pseudomonas aeruginosa biofilms to ciprofloxacin and tobramycin. Antimicrob Agents Chemother 47:317–323. doi: 10.1128/AAC.47.1.317-323.2003. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
