

CTCF boundary remodels chromatin domain and drives aberrant HOX gene transcription in acute myeloid leukemia
- PMID: 29760161
- PMCID: PMC6107877
- DOI: 10.1182/blood-2017-11-814319
CTCF boundary remodels chromatin domain and drives aberrant HOX gene transcription in acute myeloid leukemia
Abstract
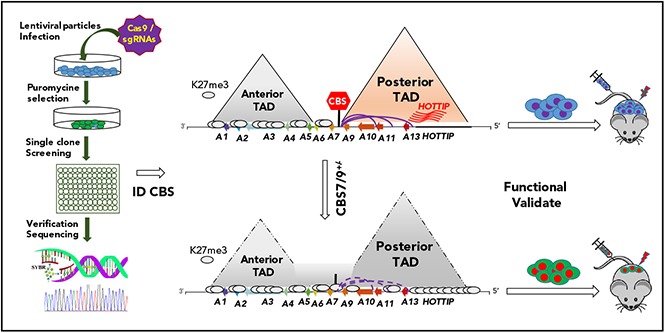
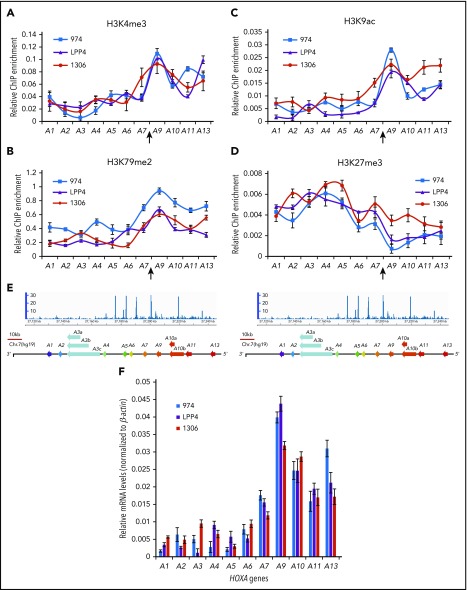
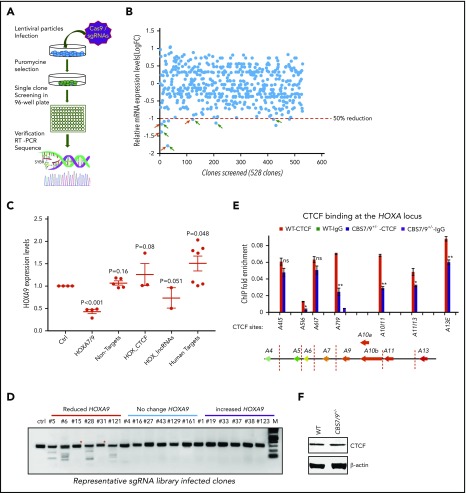
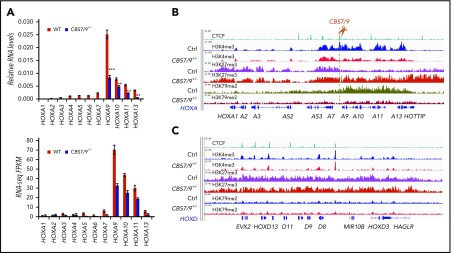
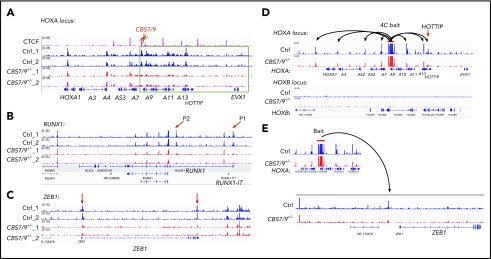
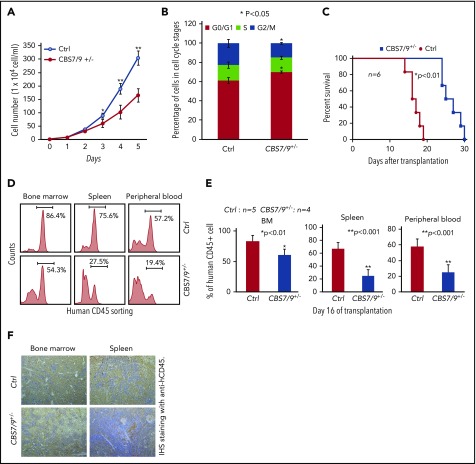
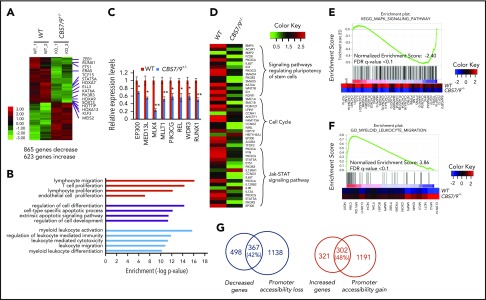
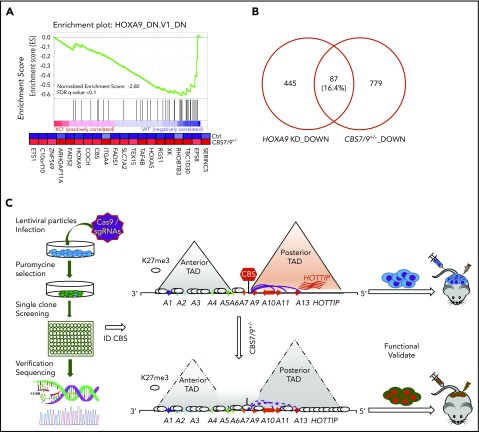
HOX gene dysregulation is a common feature of acute myeloid leukemia (AML). The molecular mechanisms underlying aberrant HOX gene expression and associated AML pathogenesis remain unclear. The nuclear protein CCCTC-binding factor (CTCF), when bound to insulator sequences, constrains temporal HOX gene-expression patterns within confined chromatin domains for normal development. Here, we used targeted pooled CRISPR-Cas9-knockout library screening to interrogate the function of CTCF boundaries in the HOX gene loci. We discovered that the CTCF binding site located between HOXA7 and HOXA9 genes (CBS7/9) is critical for establishing and maintaining aberrant HOXA9-HOXA13 gene expression in AML. Disruption of the CBS7/9 boundary resulted in spreading of repressive H3K27me3 into the posterior active HOXA chromatin domain that subsequently impaired enhancer/promoter chromatin accessibility and disrupted ectopic long-range interactions among the posterior HOXA genes. Consistent with the role of the CBS7/9 boundary in HOXA locus chromatin organization, attenuation of the CBS7/9 boundary function reduced posterior HOXA gene expression and altered myeloid-specific transcriptome profiles important for pathogenesis of myeloid malignancies. Furthermore, heterozygous deletion of the CBS7/9 chromatin boundary in the HOXA locus reduced human leukemic blast burden and enhanced survival of transplanted AML cell xenograft and patient-derived xenograft mouse models. Thus, the CTCF boundary constrains the normal gene-expression program, as well as plays a role in maintaining the oncogenic transcription program for leukemic transformation. The CTCF boundaries may serve as novel therapeutic targets for the treatment of myeloid malignancies.
© 2018 by The American Society of Hematology.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
Similar articles
-
Disruption of CTCF Boundary at HOXA Locus Promote BET Inhibitors' Therapeutic Sensitivity in Acute Myeloid Leukemia.Stem Cell Rev Rep. 2020 Dec;16(6):1280-1291. doi: 10.1007/s12015-020-10057-y. Epub 2020 Oct 14. Stem Cell Rev Rep. 2020. PMID: 33057942
-
HOX Loci Focused CRISPR/sgRNA Library Screening Identifying Critical CTCF Boundaries.J Vis Exp. 2019 Mar 31;(145):10.3791/59382. doi: 10.3791/59382. J Vis Exp. 2019. PMID: 30985763 Free PMC article.
-
Contribution of CTCF binding to transcriptional activity at the HOXA locus in NPM1-mutant AML cells.Leukemia. 2021 Feb;35(2):404-416. doi: 10.1038/s41375-020-0856-3. Epub 2020 May 12. Leukemia. 2021. PMID: 32398790 Free PMC article.
-
Therapeutic targeting in pediatric acute myeloid leukemia with aberrant HOX/MEIS1 expression.Eur J Med Genet. 2023 Dec;66(12):104869. doi: 10.1016/j.ejmg.2023.104869. Epub 2023 Oct 29. Eur J Med Genet. 2023. PMID: 38174649 Free PMC article. Review.
-
HOX gene regulation in acute myeloid leukemia: CDX marks the spot?Cell Cycle. 2007 Sep 15;6(18):2241-5. doi: 10.4161/cc.6.18.4656. Epub 2007 Jun 29. Cell Cycle. 2007. PMID: 17881901 Review.
Cited by
-
Loss of KMT2C reprograms the epigenomic landscape in hPSCs resulting in NODAL overexpression and a failure of hemogenic endothelium specification.Epigenetics. 2022 Jan-Feb;17(2):220-238. doi: 10.1080/15592294.2021.1954780. Epub 2021 Jul 24. Epigenetics. 2022. PMID: 34304711 Free PMC article.
-
Determinants and role of chromatin organization in acute leukemia.Leukemia. 2020 Oct;34(10):2561-2575. doi: 10.1038/s41375-020-0981-z. Epub 2020 Jul 20. Leukemia. 2020. PMID: 32690881 Free PMC article. Review.
-
The insulator functions of the Drosophila polydactyl C2H2 zinc finger protein CTCF: Necessity versus sufficiency.Sci Adv. 2020 Mar 25;6(13):eaaz3152. doi: 10.1126/sciadv.aaz3152. eCollection 2020 Mar. Sci Adv. 2020. PMID: 32232161 Free PMC article.
-
Chromosome structural variation in tumorigenesis: mechanisms of formation and carcinogenesis.Epigenetics Chromatin. 2020 Nov 10;13(1):49. doi: 10.1186/s13072-020-00371-7. Epigenetics Chromatin. 2020. PMID: 33168103 Free PMC article. Review.
-
TAD-dependent sub-TAD is required for enhancer-promoter interaction enabling the β-globin transcription.FASEB J. 2024 Nov 30;38(22):e70181. doi: 10.1096/fj.202401526RR. FASEB J. 2024. PMID: 39545685 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
