

CO2-sensitive tRNA modification associated with human mitochondrial disease
- PMID: 29760464
- PMCID: PMC5951830
- DOI: 10.1038/s41467-018-04250-4
CO2-sensitive tRNA modification associated with human mitochondrial disease
Abstract
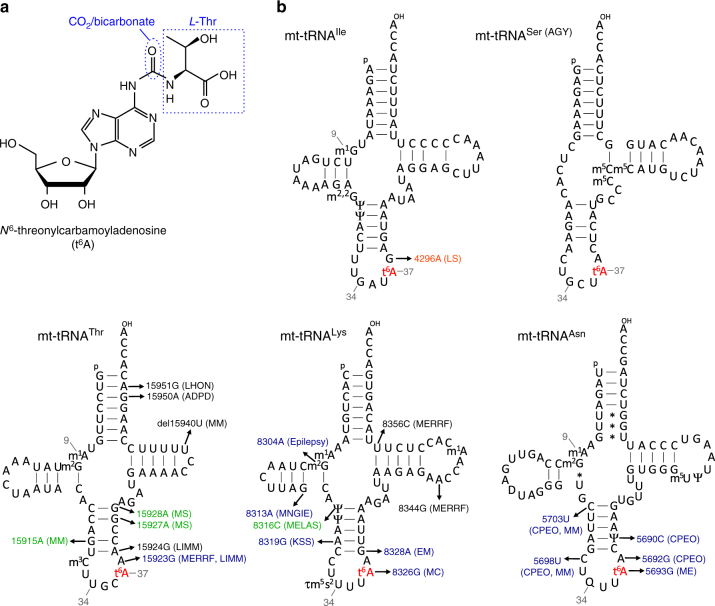
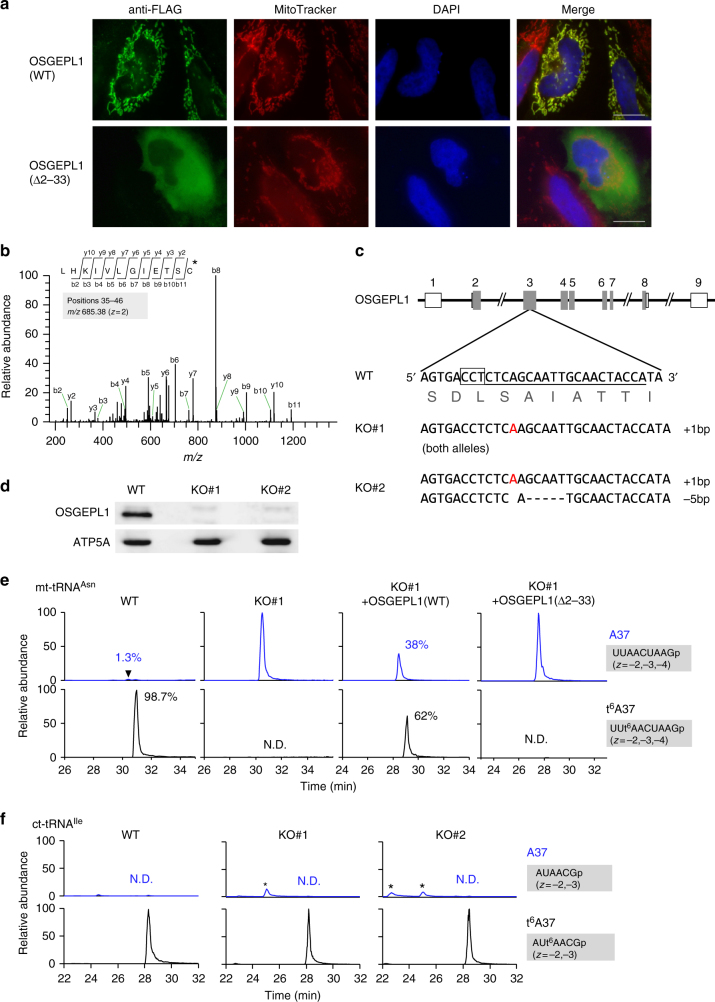
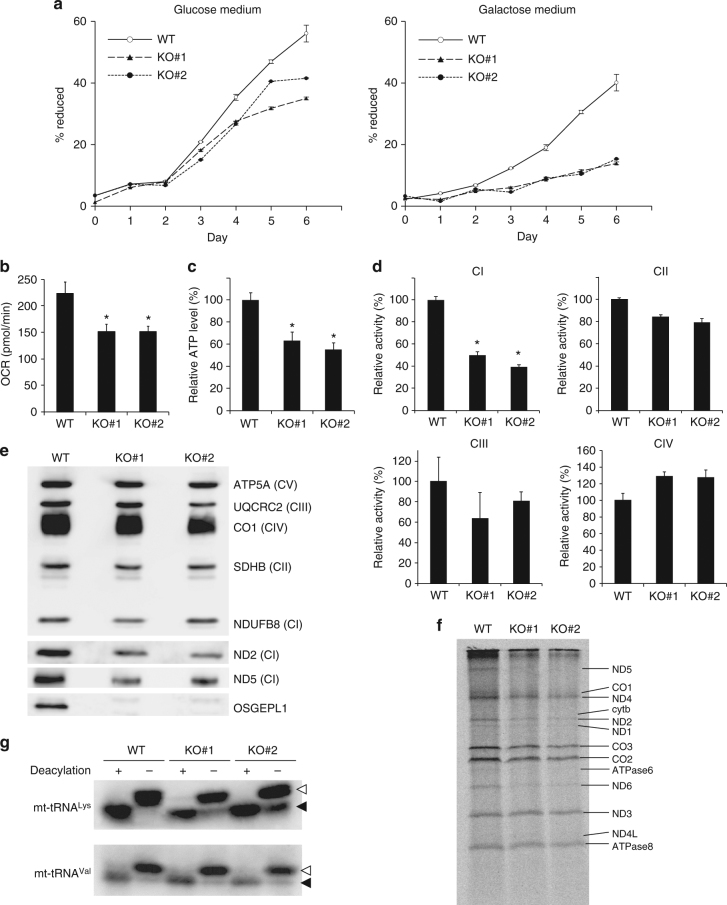
It has been generally thought that tRNA modifications are stable and static, and their frequencies are rarely regulated. N6-threonylcarbamoyladenosine (t6A) occurs at position 37 of five mitochondrial (mt-)tRNA species. We show that YRDC and OSGEPL1 are responsible for t6A37 formation, utilizing L-threonine, ATP, and CO2/bicarbonate as substrates. OSGEPL1-knockout cells exhibit respiratory defects and reduced mitochondrial translation. We find low level of t6A37 in mutant mt-tRNA isolated from the MERRF-like patient's cells, indicating that lack of t6A37 results in pathological consequences. Kinetic measurements of t6A37 formation reveal that the Km value of CO2/bicarbonate is extremely high (31 mM), suggesting that CO2/bicarbonate is a rate-limiting factor for t6A37 formation. Consistent with this, we observe a low frequency of t6A37 in mt-tRNAs isolated from human cells cultured without bicarbonate. These findings indicate that t6A37 is regulated by sensing intracellular CO2/bicarbonate concentration, implying that mitochondrial translation is modulated in a codon-specific manner under physiological conditions.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Suzuki, T. in Fine-Tuning of RNA Functions by Modification and Editing Vol. 12 (ed. Henri, G.) 23–69 (Springer-Verlag, Berlin-Heidelberg, 2005).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
