

Silver Nanoparticles: Two-Faced Neuronal Differentiation-Inducing Material in Neuroblastoma (SH-SY5Y) Cells
- PMID: 29762523
- PMCID: PMC5983825
- DOI: 10.3390/ijms19051470
Silver Nanoparticles: Two-Faced Neuronal Differentiation-Inducing Material in Neuroblastoma (SH-SY5Y) Cells
Abstract
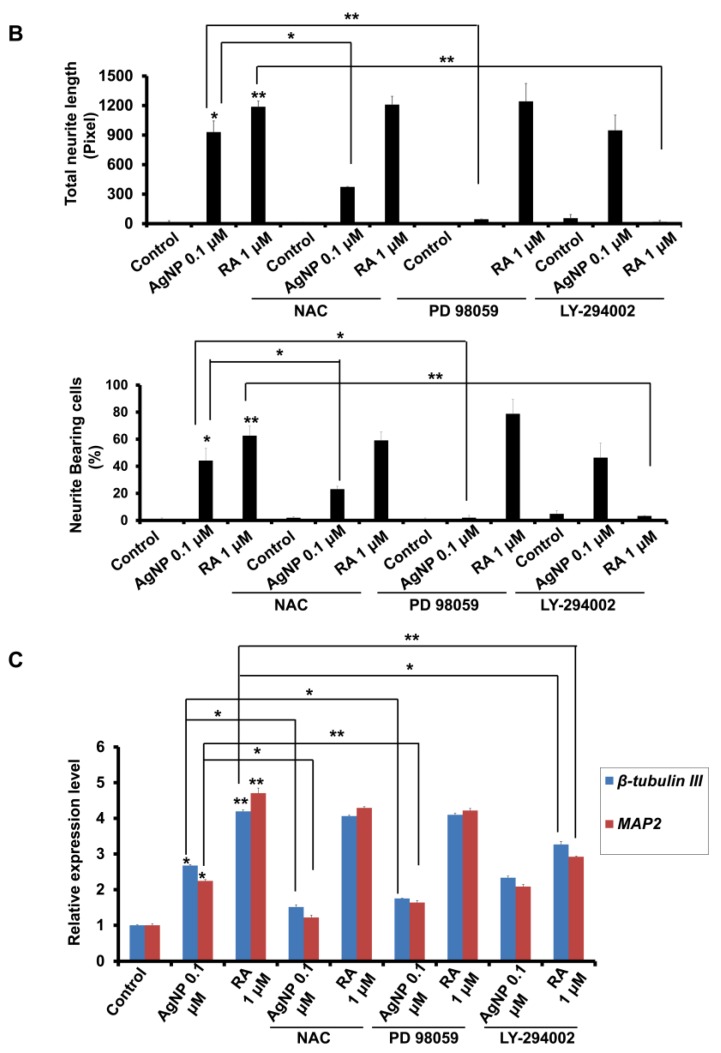
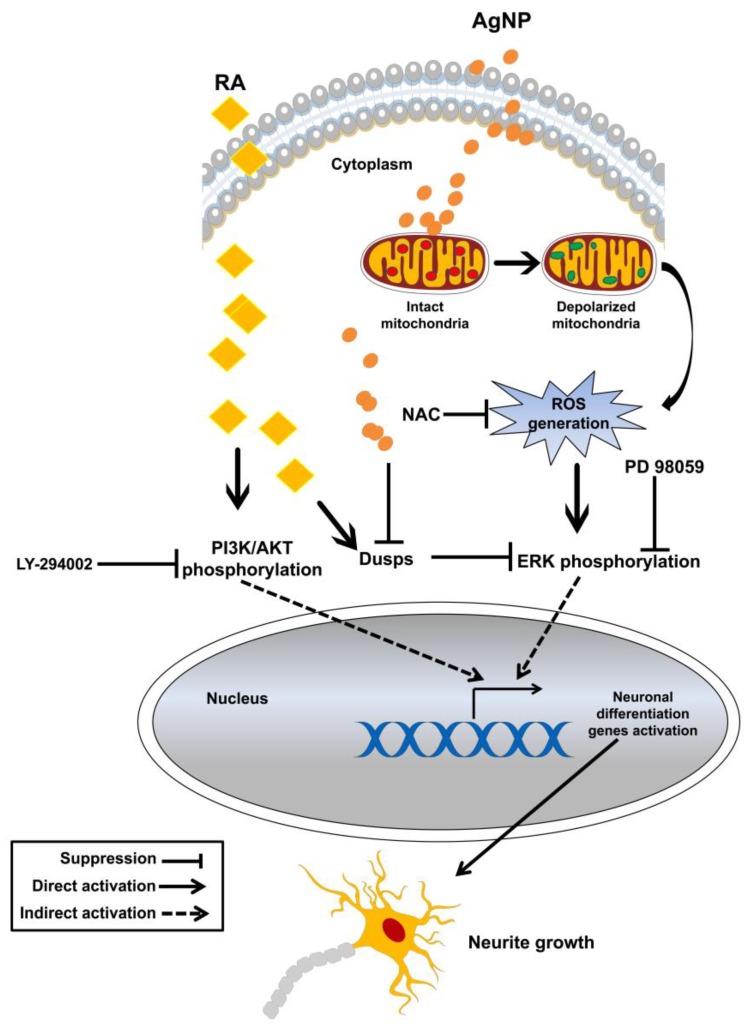
We have previously demonstrated the potential of biologically synthesized silver nanoparticles (AgNP) in the induction of neuronal differentiation of human neuroblastoma, SH-SY5Y cells; we aimed herein to unveil its molecular mechanism in comparison to the well-known neuronal differentiation-inducing agent, all-trans-retinoic acid (RA). AgNP-treated SH-SY5Y cells showed significantly higher reactive oxygen species (ROS) generation, stronger mitochondrial membrane depolarization, lower dual-specificity phosphatase expression, higher extracellular-signal-regulated kinase (ERK) phosphorylation, lower AKT phosphorylation, and lower expression of the genes encoding the antioxidant enzymes than RA-treated cells. Notably, pretreatment with N-acetyl-l-cysteine significantly abolished AgNP-induced neuronal differentiation, but not in that induced by RA. ERK inhibition, but not AKT inhibition, suppresses neurite growth that is induced by AgNP. Taken together, our results uncover the pivotal contribution of ROS in the AgNP-induced neuronal differentiation mechanism, which is different from that of RA. However, the negative consequence of AgNP-induced neurite growth may be high ROS generation and the downregulation of the expression of the genes encoding the antioxidant enzymes, which prompts the future consideration and an in-depth study of the application of AgNP-differentiated cells in neurodegenerative disease therapy.
Keywords: differentiation; kinase; mitochondria; neuroblastoma; phosphatase; reactive oxygen species; silver nanoparticles.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
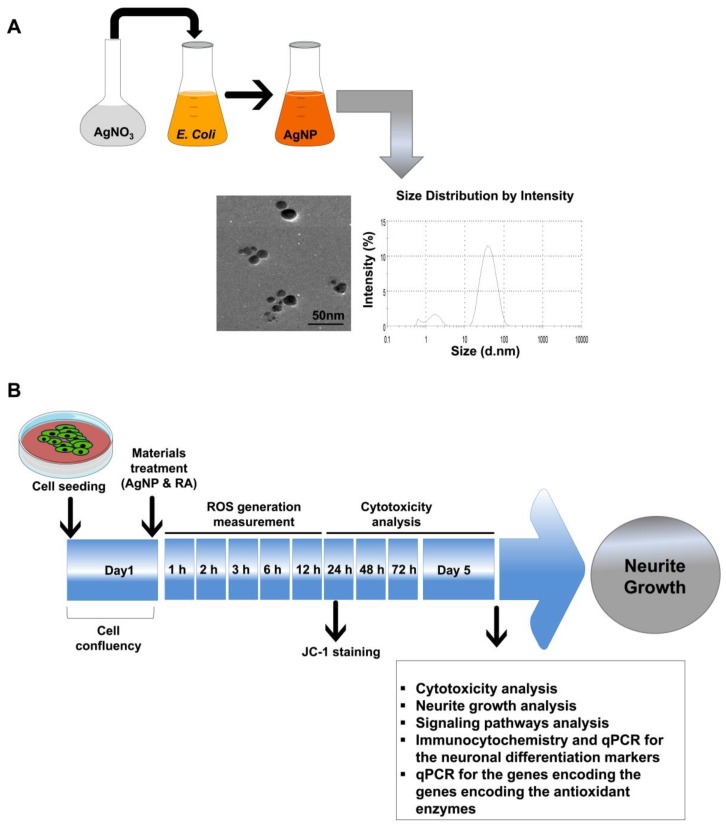
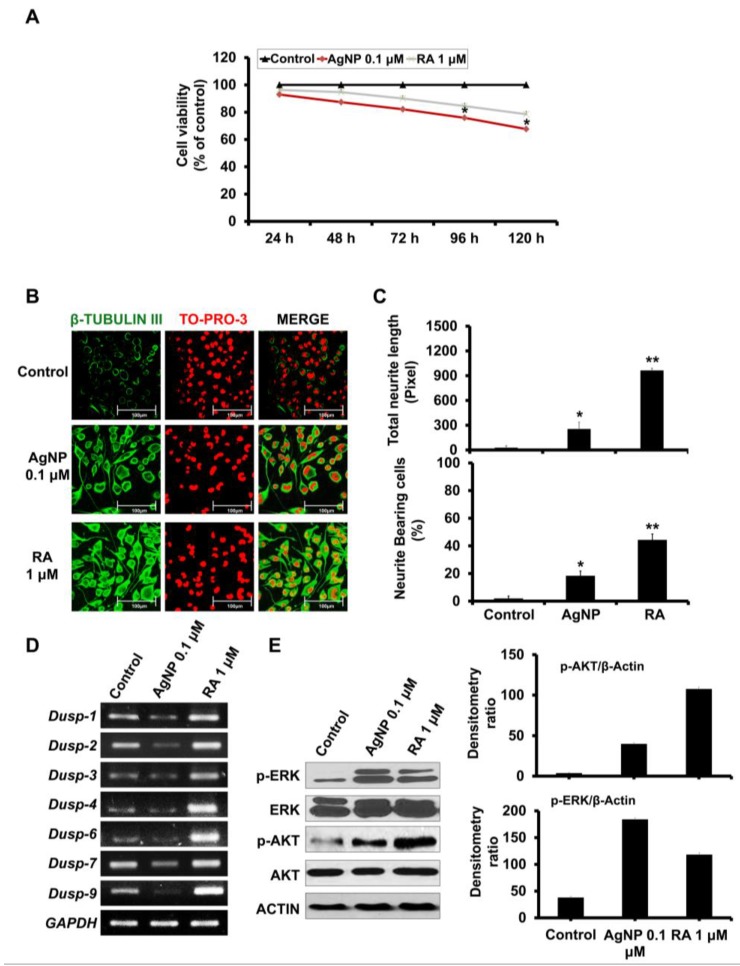
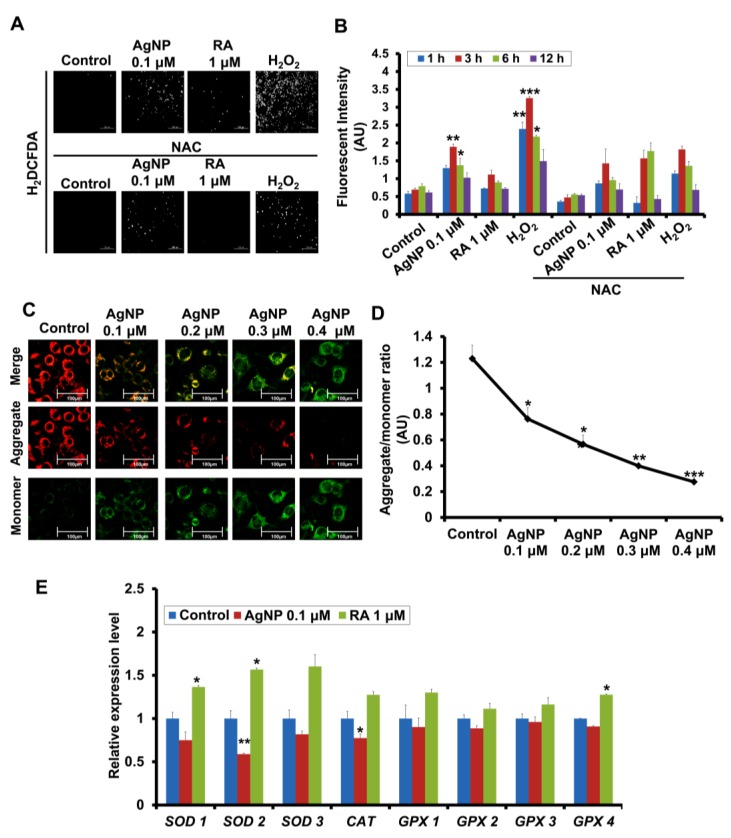
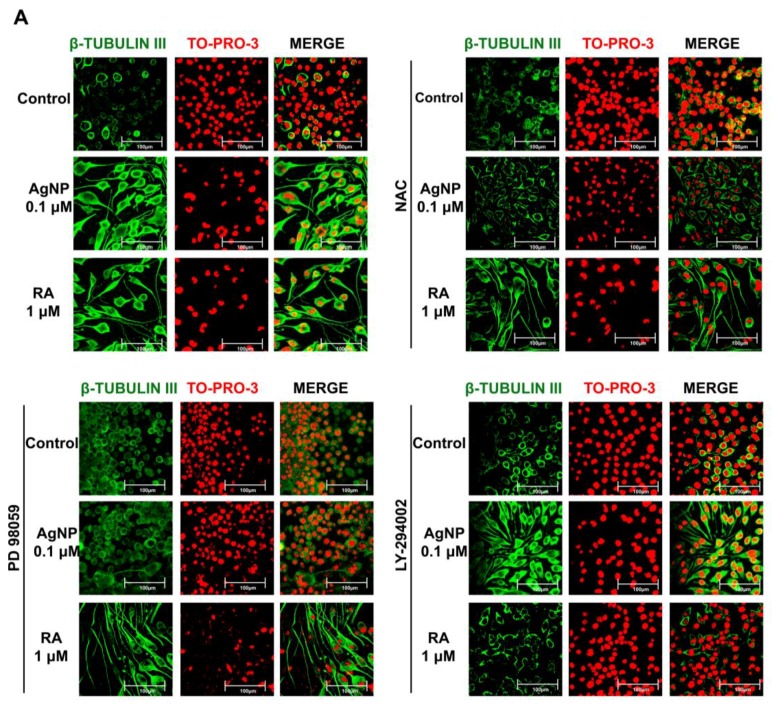
References
-
- Lopes F.M., Schröder R., da Frota Júnior M.L.C., Zanotto-Filho A., Müller C.B., Pires A.S., Meurer R.T., Colpo G.D., Gelain D.P., Kapczinski F., et al. Comparison between proliferative and neuron-like SH-SY5Y cells as an in vitro model for parkinson disease studies. Brain Res. 2010;1337:85–94. doi: 10.1016/j.brainres.2010.03.102. - DOI - PubMed
-
- Krishna A., Biryukov M., Trefois C., Antony P.M., Hussong R., Lin J., Heinäniemi M., Glusman G., Köglsberger S., Boyd O., et al. Systems genomics evaluation of the SH-SY5Y neuroblastoma cell line as a model for parkinson’s disease. BMC Genom. 2014;15:1154. doi: 10.1186/1471-2164-15-1154. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
