

Functional significance of co-occurring mutations in PIK3CA and MAP3K1 in breast cancer
- PMID: 29765551
- PMCID: PMC5940413
- DOI: 10.18632/oncotarget.25118
Functional significance of co-occurring mutations in PIK3CA and MAP3K1 in breast cancer
Abstract
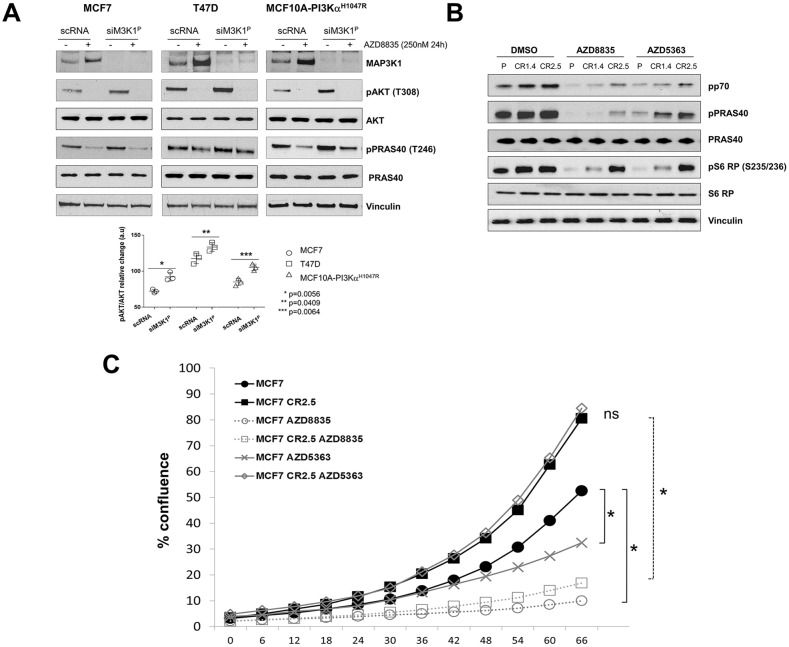
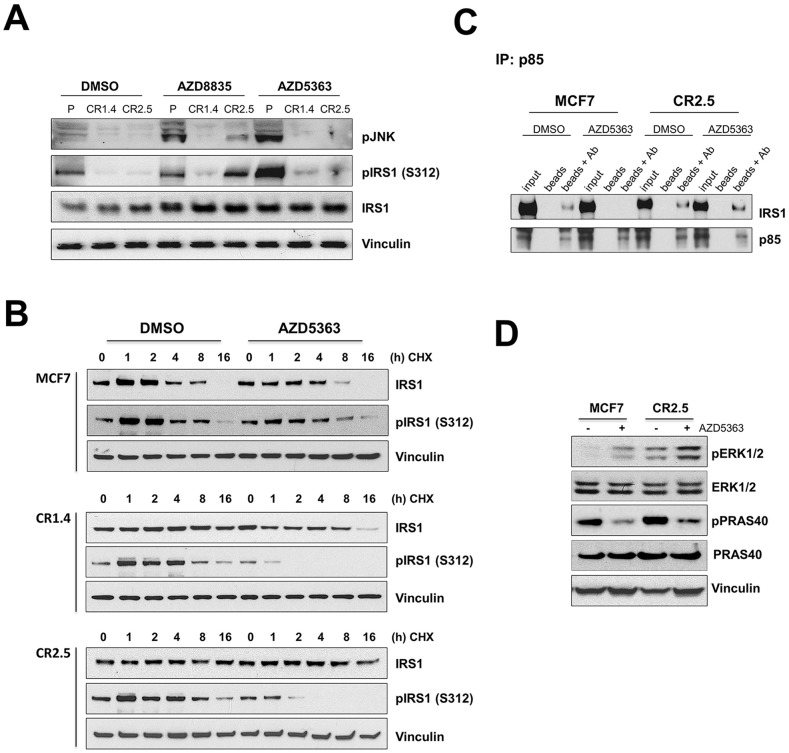
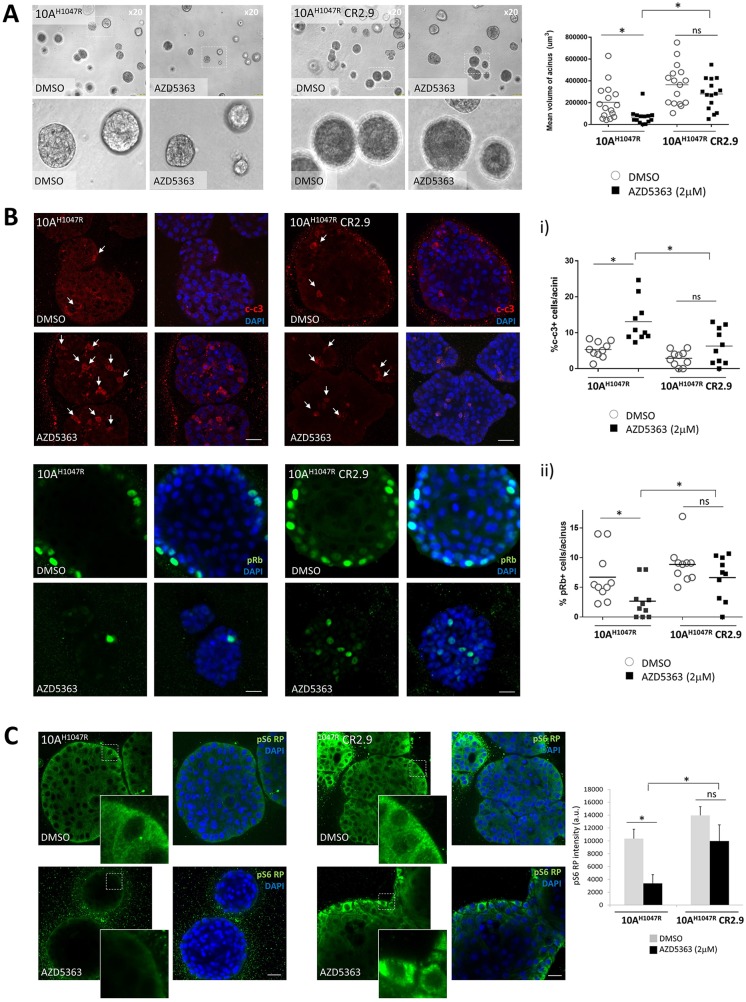
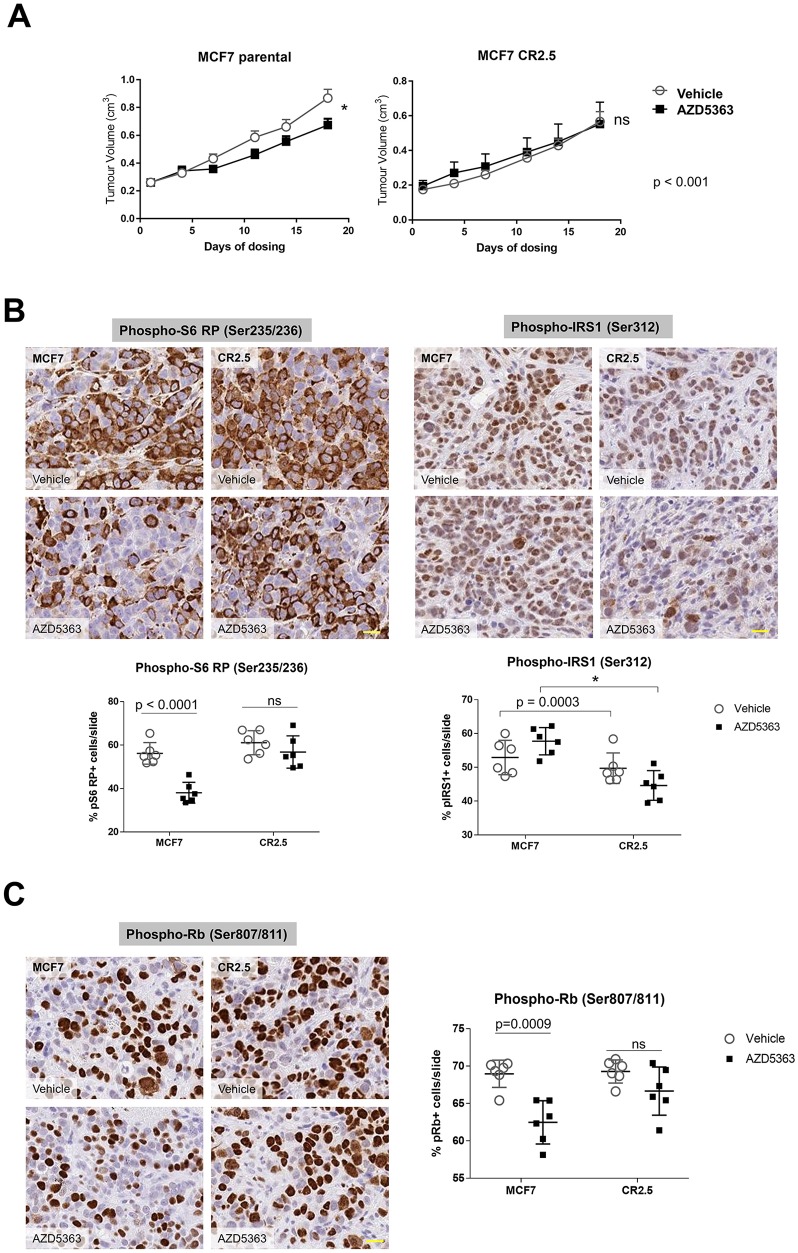
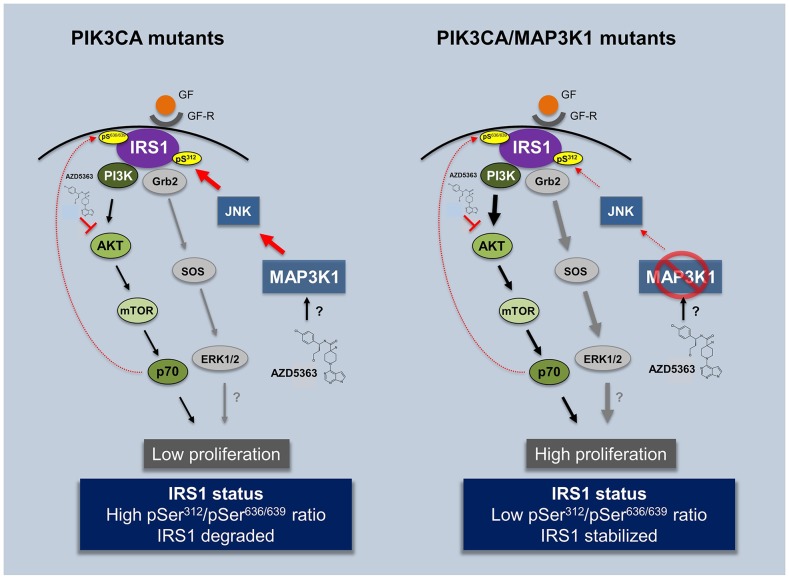
The PI3Kα signaling pathway is frequently hyper-activated in breast cancer (BrCa), as a result of mutations/amplifications in oncogenes (e.g. HER2), decreased function in tumor suppressors (e.g. PTEN) or activating mutations in key components of the pathway. In particular, activating mutations of PIK3CA (~45%) are frequently found in luminal A BrCa samples. Genomic studies have uncovered inactivating mutations in MAP3K1 (13-20%) and MAP2K4 (~8%), two upstream kinases of the JNK apoptotic pathway in luminal A BrCa samples. Further, simultaneous mutation of PIK3CA and MAP3K1 are found in ~11% of mutant PIK3CA tumors. How these two alterations may cooperate to elicit tumorigenesis and impact the sensitivity to PI3K and AKT inhibitors is currently unknown. Using CRISPR gene editing we have genetically disrupted MAP3K1 expression in mutant PIK3CA cell lines to specifically create in vitro models reflecting the mutational status of PIK3CA and MAP3K1 in BrCa patients. MAP3K1 deficient cell lines exhibited ~2.4-fold increased proliferation rate and decreased sensitivity to PI3Kα/δ(AZD8835) and AKT (AZD5363) inhibitors (~2.61 and ~5.23-fold IC50 increases, respectively) compared with parental control cell lines. In addition, mechanistic analysis revealed that MAP3K1 disruption enhances AKT phosphorylation and downstream signaling and reduces sensitivity to AZD5363-mediated pathway inhibition. This appears to be a consequence of deficient MAP3K1-JNK signaling increasing IRS1 stability and therefore promoting IRS1 binding to p85, resulting in enhanced PI3Kα activity. Using 3D-MCF10A-PI3KαH1047R models, we found that MAP3K1 depletion increased overall acinar volume and counteracted AZD5363-mediated reduction of acinar growth due to enhanced proliferation and reduced apoptosis. Furthermore, in vivo efficacy studies revealed that MAP3K1-deficient MCF7 tumors were less sensitive to AKT inhibitor treatment, compared with parental MCF7 tumors. Our study provides mechanistic and in vivo evidence indicating a role for MAP3K1 as a tumor suppressor gene at least in the context of PIK3CA-mutant backgrounds. Further, our work predicts that MAP3K1 mutational status may be considered as a predictive biomarker for efficacy in PI3K pathway inhibitor trials.
Keywords: CRISPR gene editing; MAP3K1; PIK3CA; breast cancer; luminal A.
Conflict of interest statement
CONFLICTS OF INTEREST All authors are current or former employees of AstraZeneca. K. Hudson and F Cruzalegui have ownership interest (including patents) and are AstraZeneca shareholders. No potential conflicts of interest were disclosed by the other authors.
Figures
References
-
- Zardavas D, Irrthum A, Swanton C, Piccart M. Clinical management of breast cancer heterogeneity. Nat Rev Clin Oncol. 2015;12:381–94. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
