

Does partial titin degradation affect sarcomere length nonuniformities and force in active and passive myofibrils?
- PMID: 29768046
- PMCID: PMC6171042
- DOI: 10.1152/ajpcell.00183.2017
Does partial titin degradation affect sarcomere length nonuniformities and force in active and passive myofibrils?
Abstract
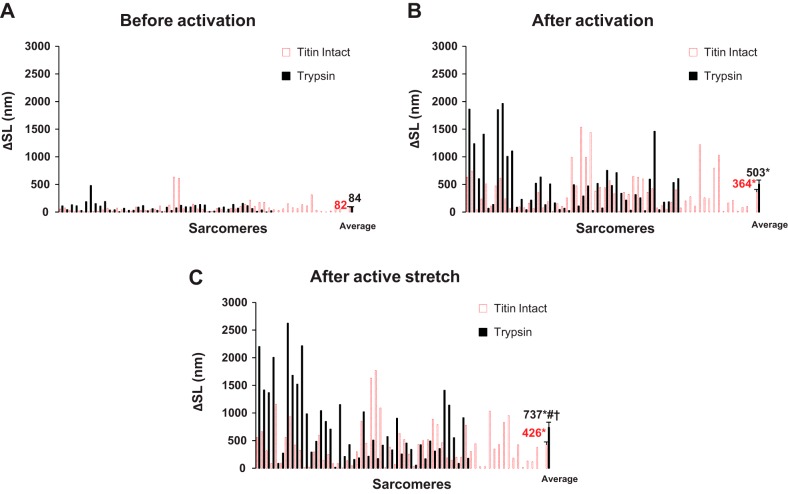
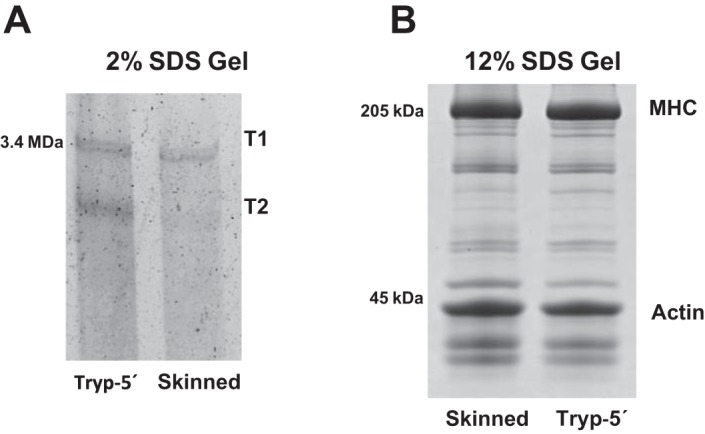
The aim of this study was to determine the role of titin in preventing the development of sarcomere length nonuniformities following activation and after active and passive stretch by determining the effect of partial titin degradation on sarcomere length nonuniformities and force in passive and active myofibrils. Selective partial titin degradation was performed using a low dose of trypsin. Myofibrils were set at a sarcomere length of 2.4 µm and then passively stretched to sarcomere lengths of 3.4 and 4.4 µm. In the active condition, myofibrils were set at a sarcomere length of 2.8 µm, activated, and actively stretched by 1 µm/sarcomere. The extent of sarcomere length nonuniformities was calculated for each sarcomere as the absolute difference between sarcomere length and the mean sarcomere length of the myofibril. Our main finding is that partial titin degradation does not increase sarcomere length nonuniformities after passive stretch and activation compared with when titin is intact but increases the extent of sarcomere length nonuniformities after active stretch. Furthermore, when titin was partially degraded, active and passive stresses were substantially reduced. These results suggest that titin plays a crucial role in actively stretched myofibrils and is likely involved in active and passive force production.
Keywords: Z-disk; active stretch; force regulation; passive stretch; sarcomere stability.
Figures






References
-
- Ait-Mou Y, Hsu K, Farman GP, Kumar M, Greaser ML, Irving TC, de Tombe PP. Titin strain contributes to the Frank-Starling law of the heart by structural rearrangements of both thin- and thick-filament proteins. Proc Natl Acad Sci USA 113: 2306–2311, 2016. doi: 10.1073/pnas.1516732113. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources