

Influenza Virus Mounts a Two-Pronged Attack on Host RNA Polymerase II Transcription
- PMID: 29768209
- PMCID: PMC5972227
- DOI: 10.1016/j.celrep.2018.04.047
Influenza Virus Mounts a Two-Pronged Attack on Host RNA Polymerase II Transcription
Abstract
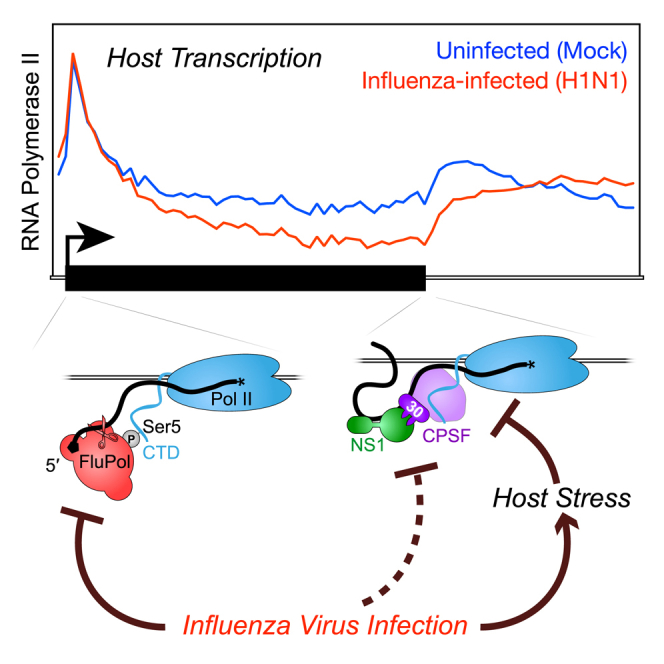
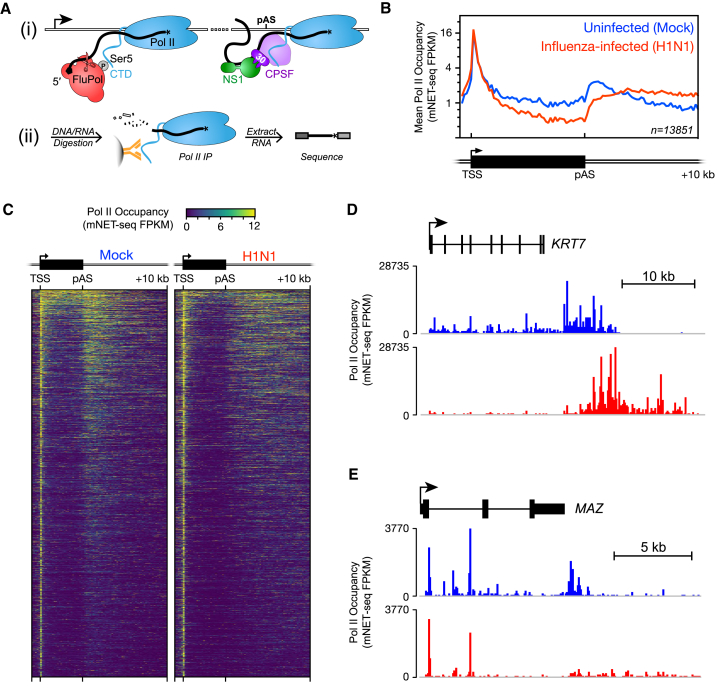
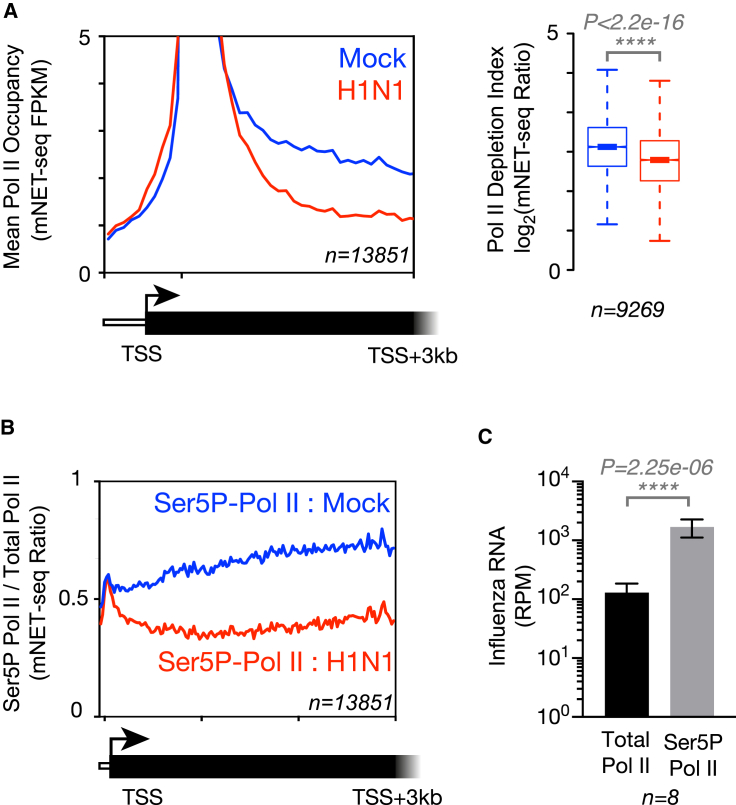
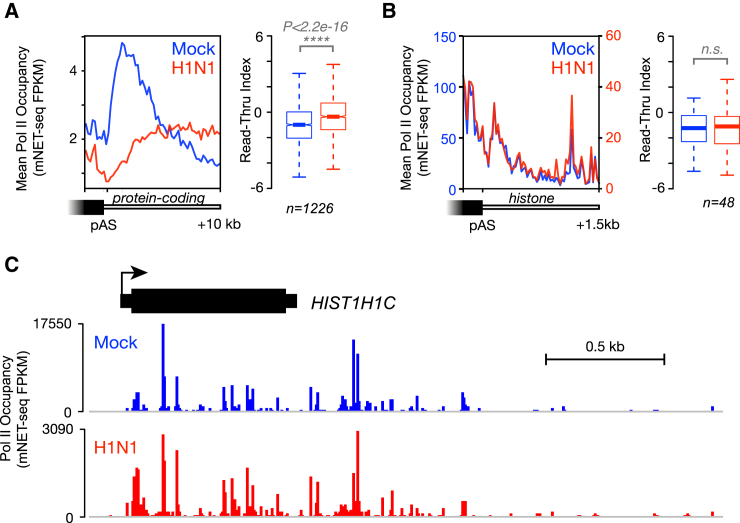
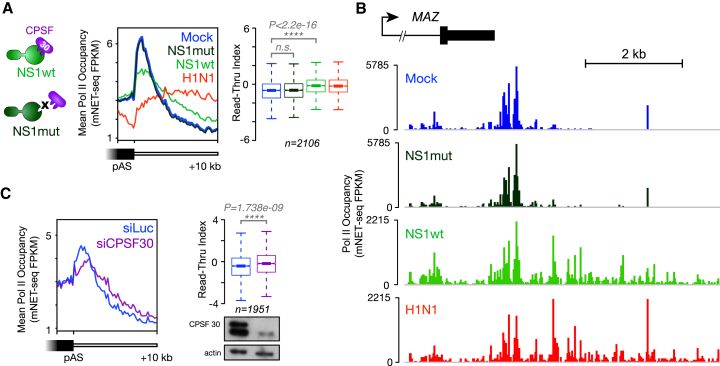
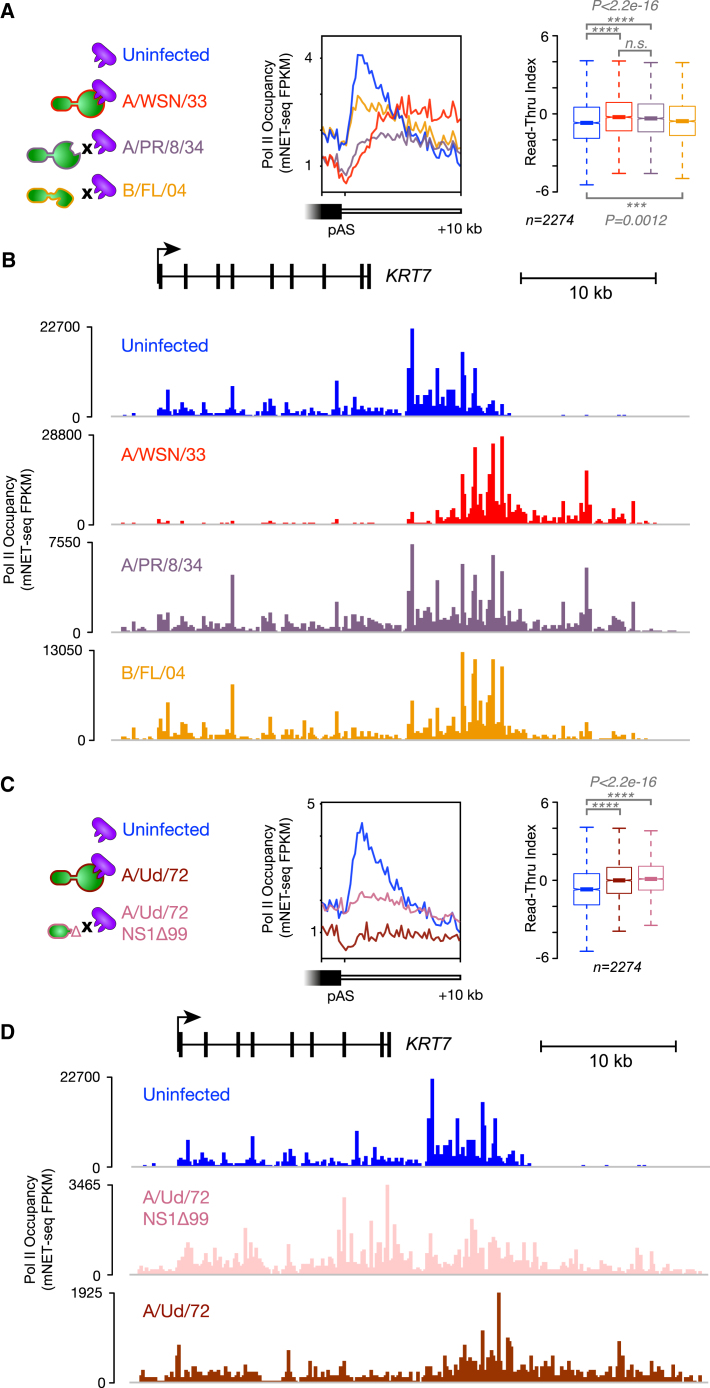
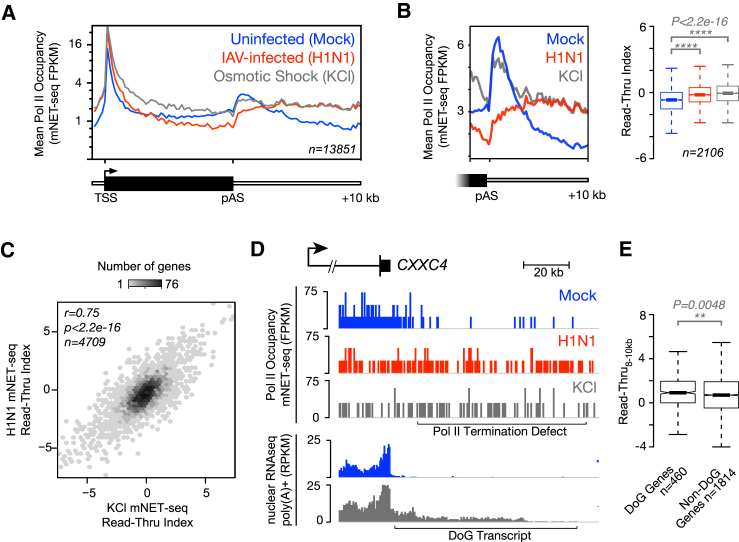
Influenza virus intimately associates with host RNA polymerase II (Pol II) and mRNA processing machinery. Here, we use mammalian native elongating transcript sequencing (mNET-seq) to examine Pol II behavior during viral infection. We show that influenza virus executes a two-pronged attack on host transcription. First, viral infection causes decreased Pol II gene occupancy downstream of transcription start sites. Second, virus-induced cellular stress leads to a catastrophic failure of Pol II termination at poly(A) sites, with transcription often continuing for tens of kilobases. Defective Pol II termination occurs independently of the ability of the viral NS1 protein to interfere with host mRNA processing. Instead, this termination defect is a common effect of diverse cellular stresses and underlies the production of previously reported downstream-of-gene transcripts (DoGs). Our work has implications for understanding not only host-virus interactions but also fundamental aspects of mammalian transcription.
Keywords: DoGs; RNA polymerase II; downstream-of-gene transcripts; influenza; transcription; transcription termination; virus-induced host shutoff.
Copyright © 2018 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Brannan K., Kim H., Erickson B., Glover-Cutter K., Kim S., Fong N., Kiemele L., Hansen K., Davis R., Lykke-Andersen J., Bentley D.L. mRNA decapping factors and the exonuclease Xrn2 function in widespread premature termination of RNA polymerase II transcription. Mol. Cell. 2012;46:311–324. - PMC - PubMed
-
- Buratowski S. Connections between mRNA 3′ end processing and transcription termination. Curr. Opin. Cell Biol. 2005;17:257–261. - PubMed
-
- Chan A.Y., Vreede F.T., Smith M., Engelhardt O.G., Fodor E. Influenza virus inhibits RNA polymerase II elongation. Virology. 2006;351:210–217. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
