

A congenic line of the C57BL/6J mouse strain that is proficient in melatonin synthesis
- PMID: 29768727
- PMCID: PMC6167157
- DOI: 10.1111/jpi.12509
A congenic line of the C57BL/6J mouse strain that is proficient in melatonin synthesis
Abstract
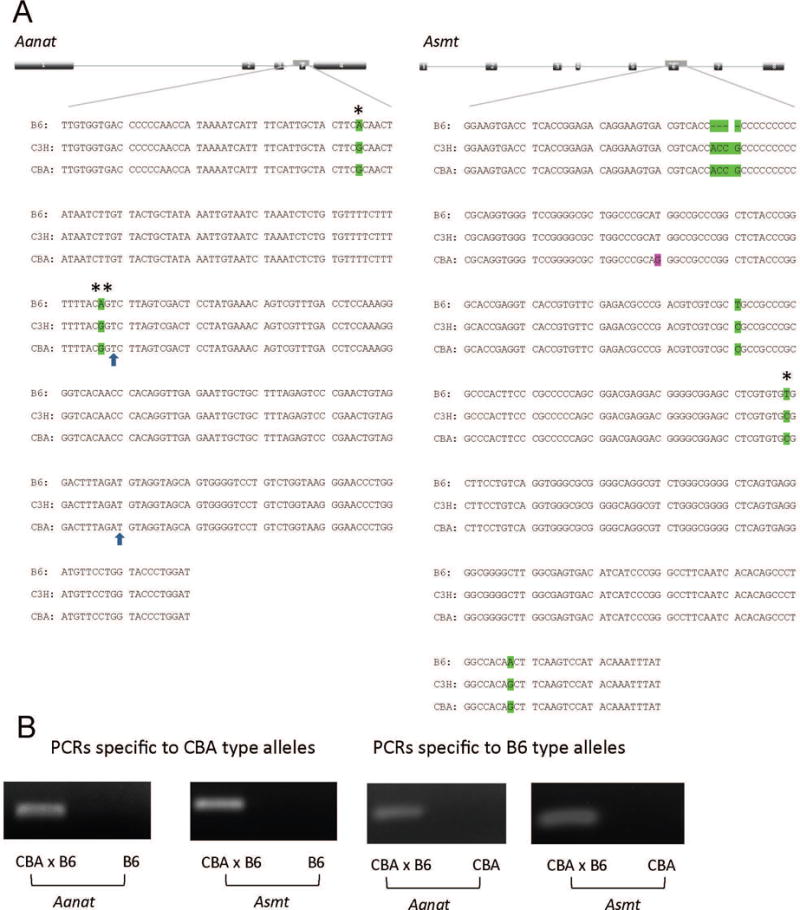
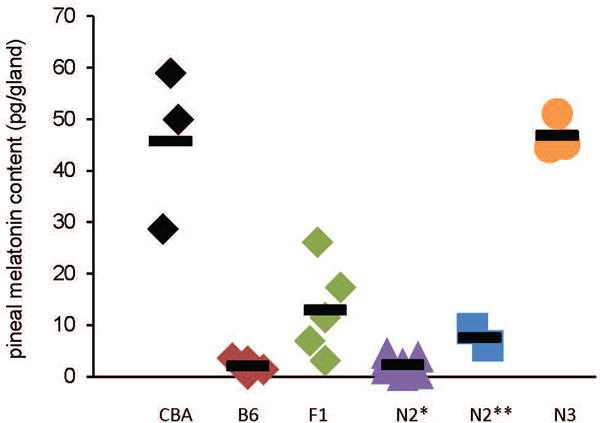
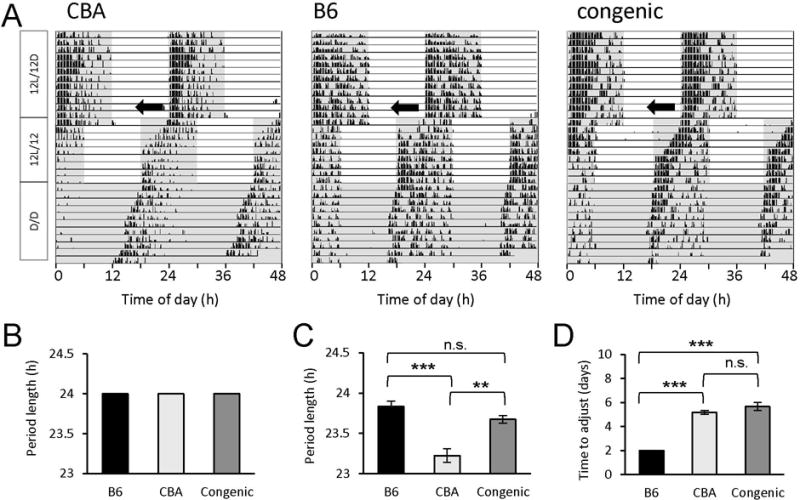
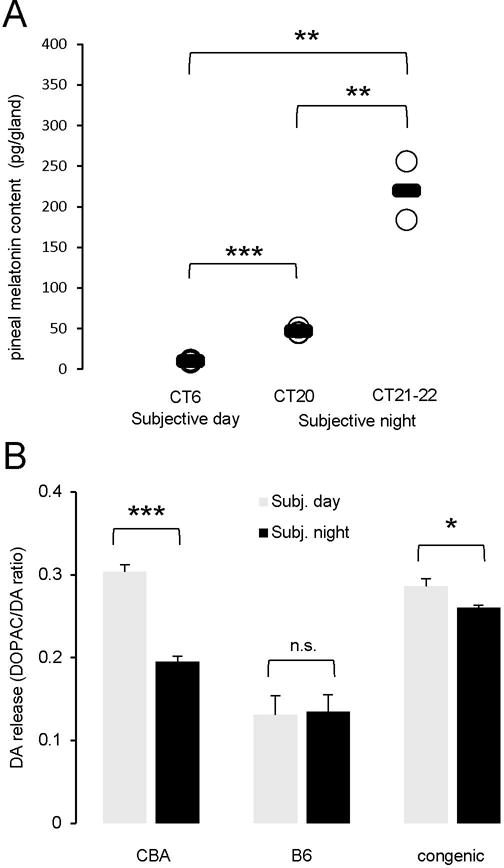
The C57BL/6J (B6) is the most common inbred mouse strain used in biomedical research in the United States. Yet, this strain is notoriously known for being deficient in the biosynthesis of melatonin, an important effector of circadian clocks in the brain and in the retina. Melatonin deficiency in this strain results from nonfunctional alleles of the genes coding 2 key enzymes of the melatonin synthesis pathway: arylalkylamine-N-acetyltransferase (Aanat) and N-acetylserotonin-O-methyltransferase (Asmt). By introducing functional alleles of the Aanat and Asmt genes from the melatonin-proficient CBA/CaJ (CBA) mouse strain to B6, we have generated a B6 congenic line that has acquired the capacity of rhythmic melatonin synthesis. In addition, the melatonin-dependent rhythm of dopamine release in the retina is restored in the B6 congenic line. Finally, we have partially characterized the Aanat and Asmt genes of the CBA strain and have identified multiple differences between CBA and B6 alleles, including single nucleotide polymorphism and deletion/insertion of DNA segments of various sizes. As an improved model organism with functional components of the melatonin synthesis pathway and melatonin-dependent circadian regulations, the new line will be useful to researchers studying melatonin physiological functions in a variety of fields including, but not limited to, circadian biology and neuroscience. In particular, the congenic line will be useful to speed up introduction of melatonin production capacity into genetically modified mouse lines of interest such as knockout lines, many of which are on B6 or mixed B6 backgrounds. The melatonin-proficient B6 congenic line will be widely distributed.
Keywords: C3Heb/FeJ; C57BL/6J; CBA/CaJ; N-acetylserotonin-O-methyltransferase; arylalkylamine-N-acetyltransferase; circadian rhythms; congenic; melatonin; retina.
© 2018 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd.
Figures
References
-
- Simonneaux V, Ribelayga C. Generation of the melatonin endocrine message in mammals: A review of the complex regulation of melatonin synthesis by norepinephrine, peptides, and other pineal transmitters. Pharmacol Rev. 2003;55:325–395. - PubMed
-
- Pevet P, Challet E. Melatonin: both master clock output and internal time-giver in the circadian clocks network. J Physiol Paris. 2011;105:170–82. - PubMed
-
- Iuvone PM, Tosini G, Pozdeyev N, et al. Circadian clocks, clock networks, arylalkylamine N-acetyltransferase, and melatonin in the retina. Prog Retin Eye Res. 2005;24:433–56. - PubMed
-
- Ebihara S, Marks T, Hudson DJ, Menaker M. Genetic control of melatonin synthesis in the pineal gland of the mouse. Science. 1986;231:491–493. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
