

Defense Mechanisms Against Acid Exposure by Dental Enamel Formation, Saliva and Pancreatic Juice Production
- PMID: 29769002
- PMCID: PMC6225347
- DOI: 10.2174/1381612824666180515125654
Defense Mechanisms Against Acid Exposure by Dental Enamel Formation, Saliva and Pancreatic Juice Production
Abstract
The pancreas, the salivary glands and the dental enamel producing ameloblasts have marked developmental, structural and functional similarities. One of the most striking similarities is their bicarbonate-rich secretory product, serving acid neutralization. An important difference between them is that while pancreatic juice and saliva are delivered into a lumen where they can be collected and analyzed, ameloblasts produce locally precipitating hydroxyapatite which cannot be easily studied. Interestingly, the ion and protein secretion by the pancreas, the salivary glands, and maturation ameloblasts are all two-step processes, of course with significant differences too. As they all have to defend against acid exposure by producing extremely large quantities of bicarbonate, the failure of this function leads to deteriorating consequences. The aim of the present review is to describe and characterize the defense mechanisms of the pancreas, the salivary glands and enamel-producing ameloblasts against acid exposure and to compare their functional capabilities to do this by producing bicarbonate.
Keywords: Salivary gland; acid; acinar cell; ameloblast; bicarbonate; defense; dental enamel; ductal cell; pancreas..
Copyright© Bentham Science Publishers; For any queries, please email at epub@benthamscience.org.
Figures
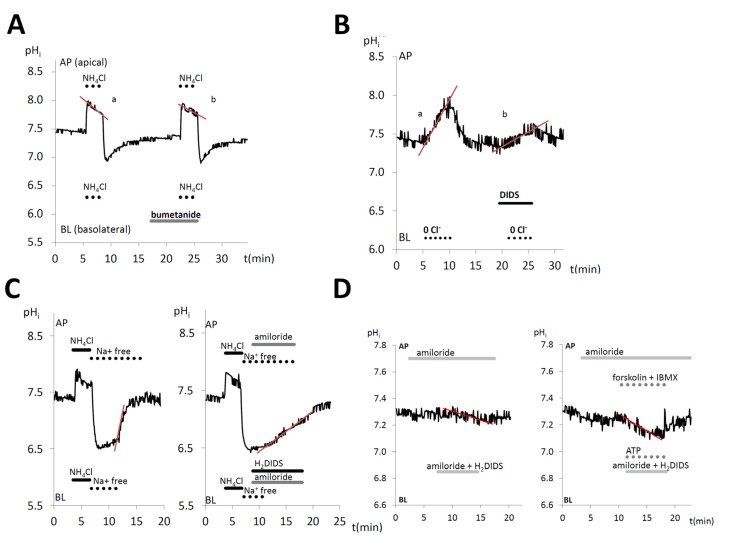
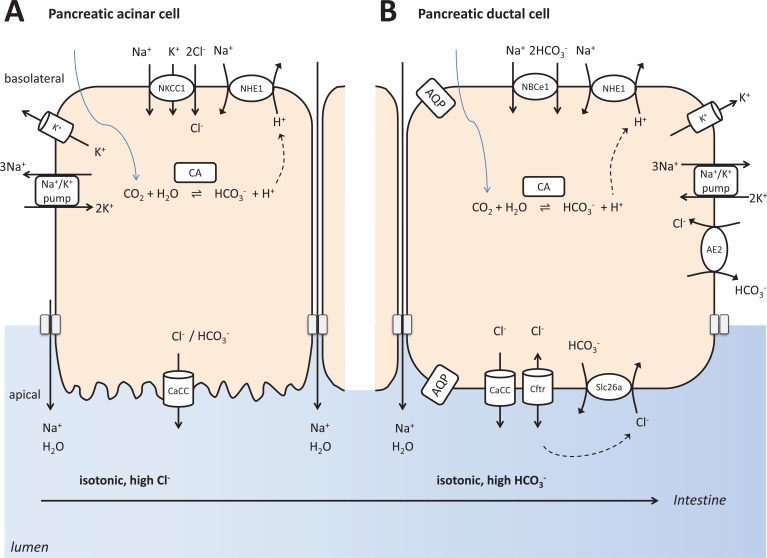
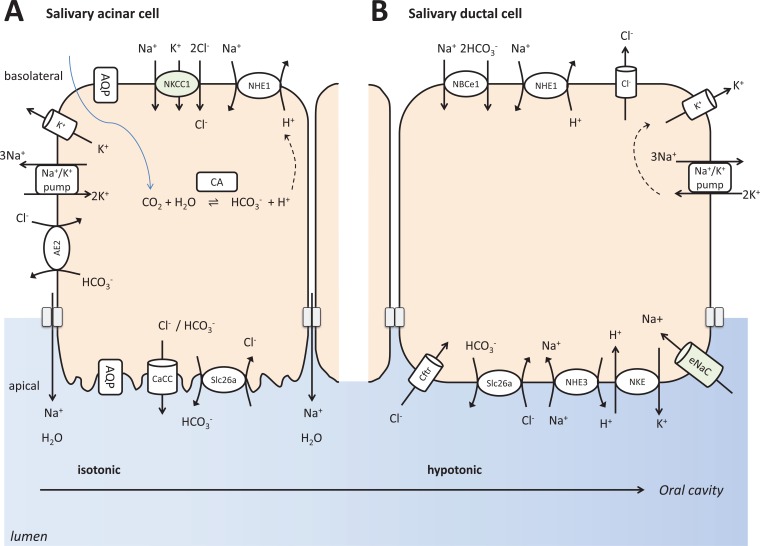
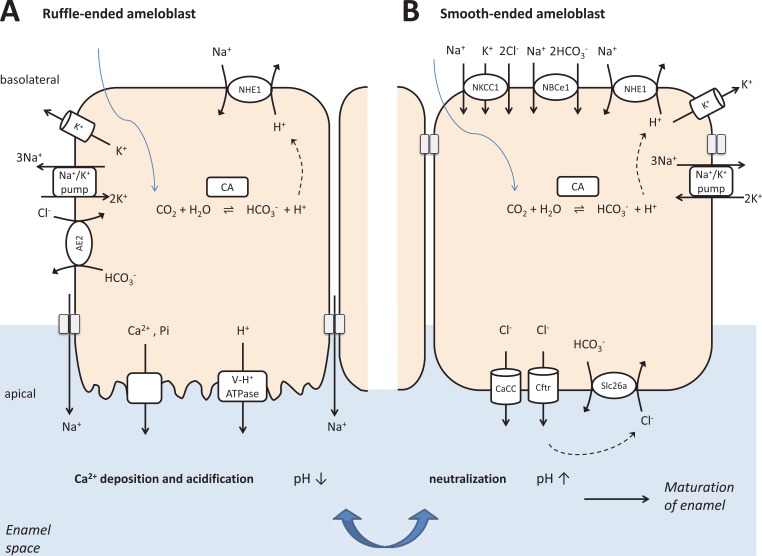
References
-
- Steward M.C., Ishiguro H., Case R.M. Mechanisms of bicarbonate secretion in the pancreatic duct. Annu. Rev. Physiol. 2005;67:377–409. - PubMed
-
- Varga G. Physiology of the salivary glands. Surgery. 2015;33:581–586.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
