

Reciprocal Negative Regulation Between Lmx1a and Lmo4 Is Required for Inner Ear Formation
- PMID: 29769265
- PMCID: PMC5990987
- DOI: 10.1523/JNEUROSCI.2484-17.2018
Reciprocal Negative Regulation Between Lmx1a and Lmo4 Is Required for Inner Ear Formation
Abstract
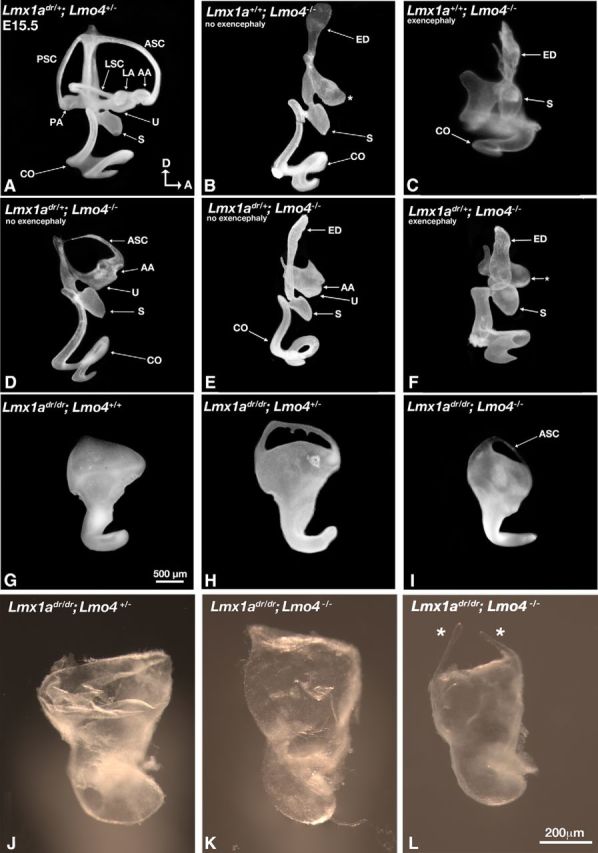
LIM-domain containing transcription factors (LIM-TFs) are conserved factors important for embryogenesis. The specificity of these factors in transcriptional regulation is conferred by the complexes that they form with other proteins such as LIM-domain-binding (Ldb) proteins and LIM-domain only (LMO) proteins. Unlike LIM-TFs, these proteins do not bind DNA directly. LMO proteins are negative regulators of LIM-TFs and function by competing with LIM-TFs for binding to Ldb's. Although the LIM-TF Lmx1a is expressed in the developing mouse hindbrain, which provides many of the extrinsic signals for inner ear formation, conditional knock-out embryos of both sexes show that the inner ear source of Lmx1a is the major contributor of ear patterning. In addition, we have found that the reciprocal interaction between Lmx1a and Lmo4 (a LMO protein within the inner ear) mediates the formation of both vestibular and auditory structures. Lmo4 negatively regulates Lmx1a to form the three sensory cristae, the anterior semicircular canal, and the shape of the utricle in the vestibule. Furthermore, this negative regulation blocks ectopic sensory formation in the cochlea. In contrast, Lmx1a negatively regulates Lmo4 in mediating epithelial resorption of the canal pouch, which gives rise to the anterior and posterior semicircular canals. We also found that Lmx1a is independently required for the formation of the endolymphatic duct and hair cells in the basal cochlear region.SIGNIFICANCE STATEMENT The mammalian inner ear is a structurally complex organ responsible for detecting sound and maintaining balance. Failure to form the intricate 3D structure of this organ properly during development most likely will result in sensory deficits on some level. Here, we provide genetic evidence that a transcription factor, Lmx1a, interacts with its negative regulator, Lmo4, to pattern various vestibular and auditory components of the mammalian inner ear. Identifying these key molecules that mediate formation of this important sensory organ will be helpful for designing strategies and therapeutics to alleviate hearing loss and balance disorders.
Keywords: cochlea; cristae; development; genetics; inner ear; semicircular canals.
Copyright © 2018 Huang et al.
Figures







Similar articles
-
Interaction with ectopic cochlear crista sensory epithelium disrupts basal cochlear sensory epithelium development in Lmx1a mutant mice.Cell Tissue Res. 2020 Jun;380(3):435-448. doi: 10.1007/s00441-019-03163-y. Epub 2020 Jan 13. Cell Tissue Res. 2020. PMID: 31932950 Free PMC article.
-
Requirement for Lmo4 in the vestibular morphogenesis of mouse inner ear.Dev Biol. 2010 Feb 1;338(1):38-49. doi: 10.1016/j.ydbio.2009.11.003. Epub 2009 Nov 10. Dev Biol. 2010. PMID: 19913004 Free PMC article.
-
Differential expression of LIM domain-only (LMO) genes in the developing mouse inner ear.Gene Expr Patterns. 2006 Oct;6(8):857-63. doi: 10.1016/j.modgep.2006.02.005. Epub 2006 Apr 4. Gene Expr Patterns. 2006. PMID: 16597514
-
LIM Homeodomain (LIM-HD) Genes and Their Co-Regulators in Developing Reproductive System and Disorders of Sex Development.Sex Dev. 2022;16(2-3):147-161. doi: 10.1159/000518323. Epub 2021 Sep 10. Sex Dev. 2022. PMID: 34518474 Review.
-
[Involvement of LMO4 in tumorigenesis associated epithelial-mesenchymal transition].Zhejiang Da Xue Xue Bao Yi Xue Ban. 2011 Jan;40(1):107-11. doi: 10.3785/j.issn.1008-9292.2011.01.019. Zhejiang Da Xue Xue Bao Yi Xue Ban. 2011. PMID: 21319383 Review. Chinese.
Cited by
-
Hear, Hear for Notch: Control of Cell Fates in the Inner Ear by Notch Signaling.Biomolecules. 2020 Feb 28;10(3):370. doi: 10.3390/biom10030370. Biomolecules. 2020. PMID: 32121147 Free PMC article. Review.
-
Development in the Mammalian Auditory System Depends on Transcription Factors.Int J Mol Sci. 2021 Apr 18;22(8):4189. doi: 10.3390/ijms22084189. Int J Mol Sci. 2021. PMID: 33919542 Free PMC article. Review.
-
Molecular mechanisms governing development of the hindbrain choroid plexus and auditory projection: A validation of the seminal observations of Wilhelm His.IBRO Neurosci Rep. 2022 Oct 3;13:306-313. doi: 10.1016/j.ibneur.2022.09.011. eCollection 2022 Dec. IBRO Neurosci Rep. 2022. PMID: 36247525 Free PMC article. Review.
-
Early Steps towards Hearing: Placodes and Sensory Development.Int J Mol Sci. 2023 Apr 10;24(8):6994. doi: 10.3390/ijms24086994. Int J Mol Sci. 2023. PMID: 37108158 Free PMC article. Review.
-
Lmx1a and Lmx1b are Redundantly Required for the Development of Multiple Components of the Mammalian Auditory System.Neuroscience. 2021 Jan 1;452:247-264. doi: 10.1016/j.neuroscience.2020.11.013. Epub 2020 Nov 24. Neuroscience. 2021. PMID: 33246067 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous