

Metarrestin, a perinucleolar compartment inhibitor, effectively suppresses metastasis
- PMID: 29769289
- PMCID: PMC6176865
- DOI: 10.1126/scitranslmed.aap8307
Metarrestin, a perinucleolar compartment inhibitor, effectively suppresses metastasis
Abstract
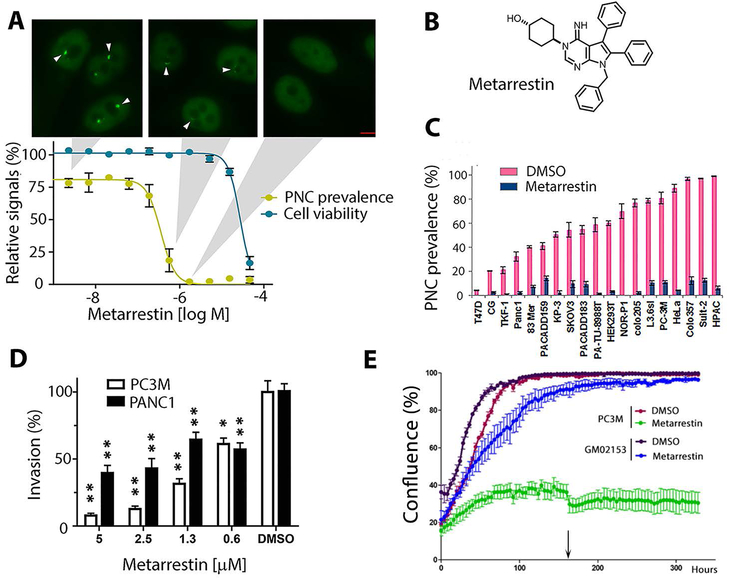
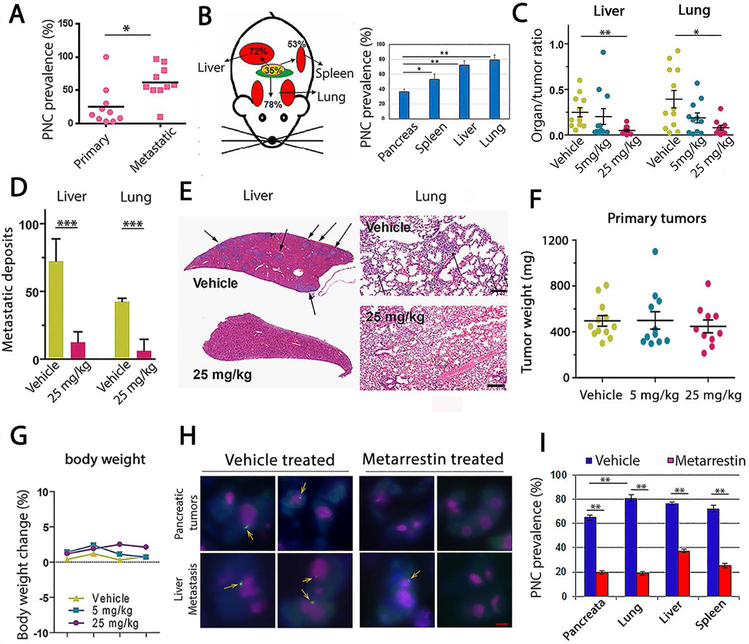
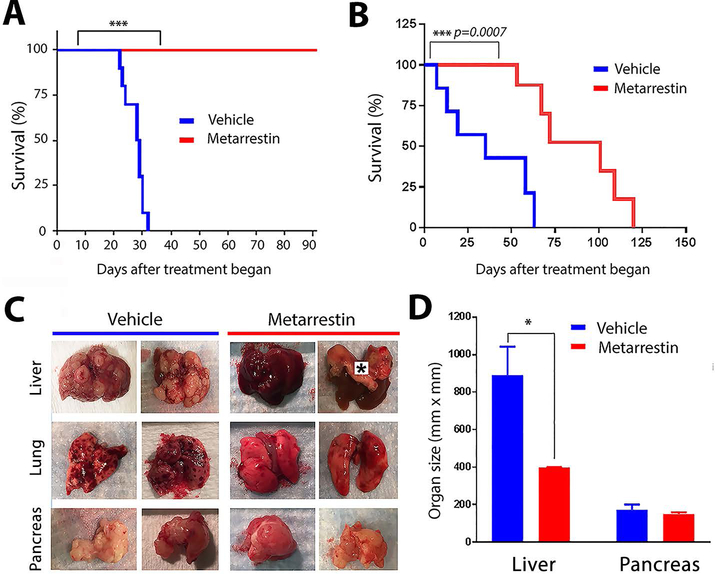
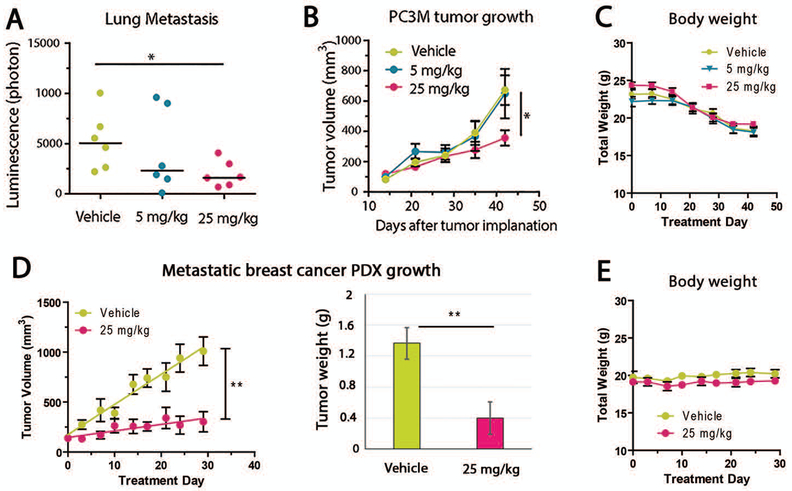
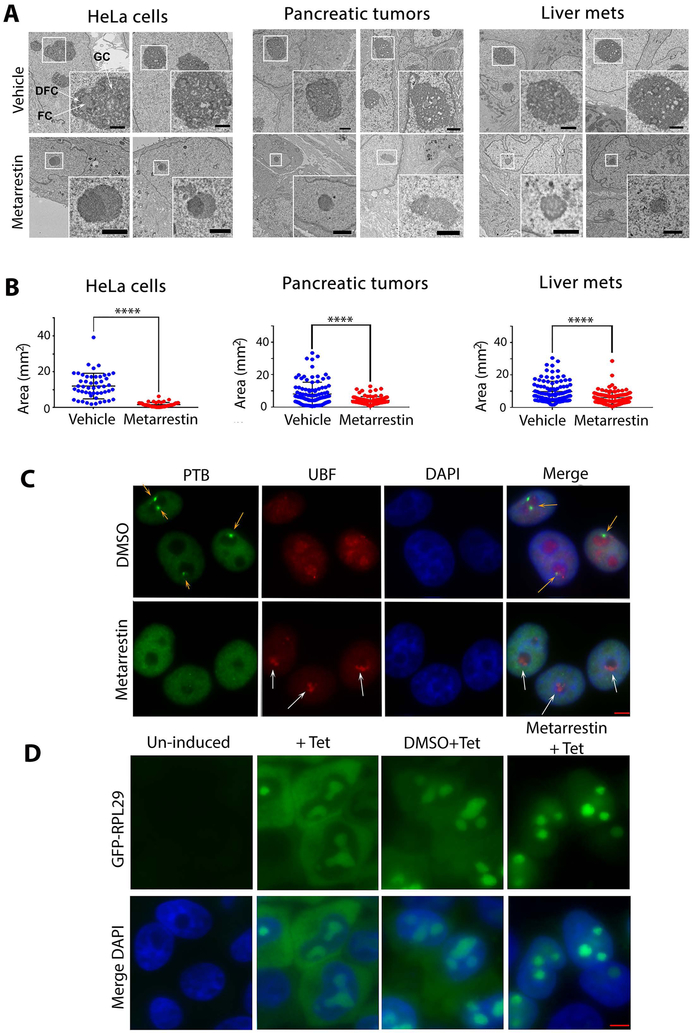
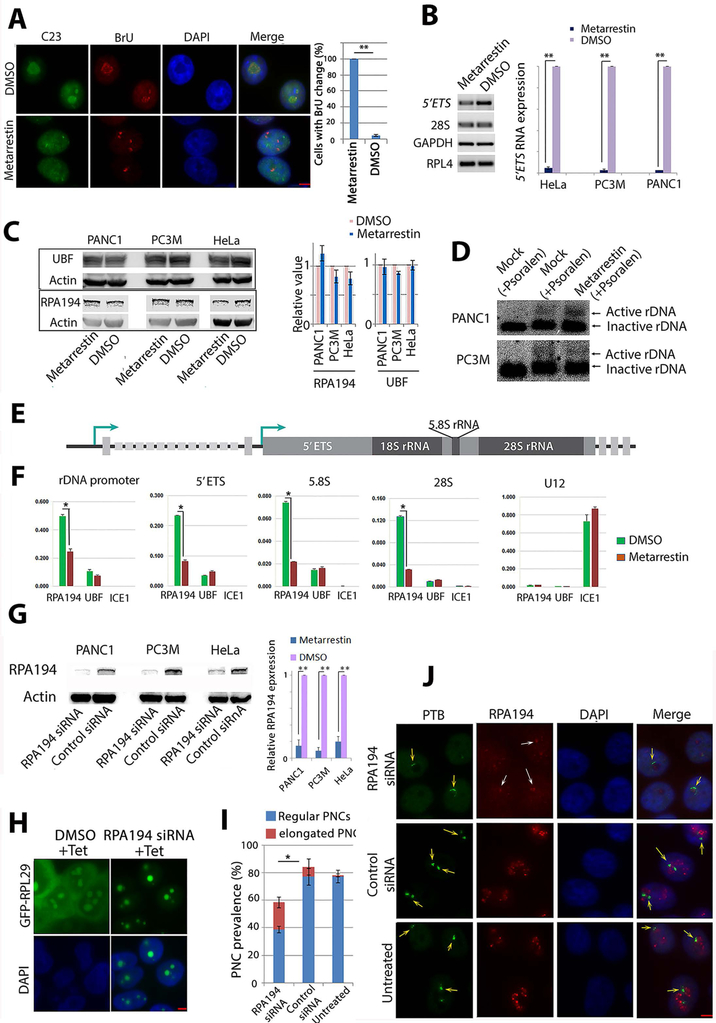
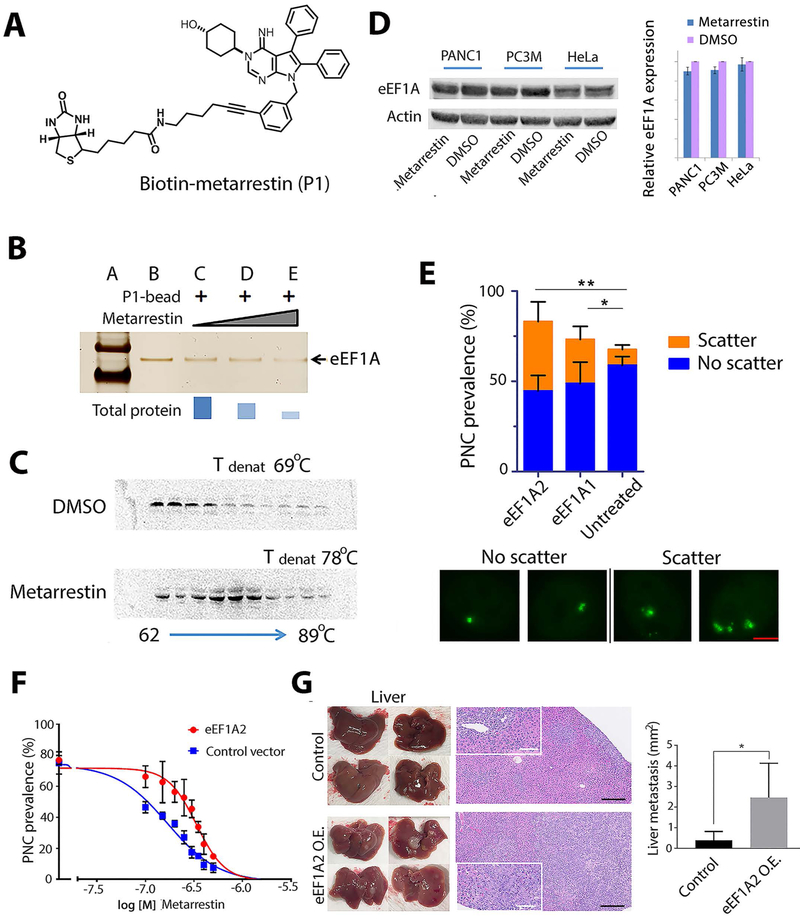
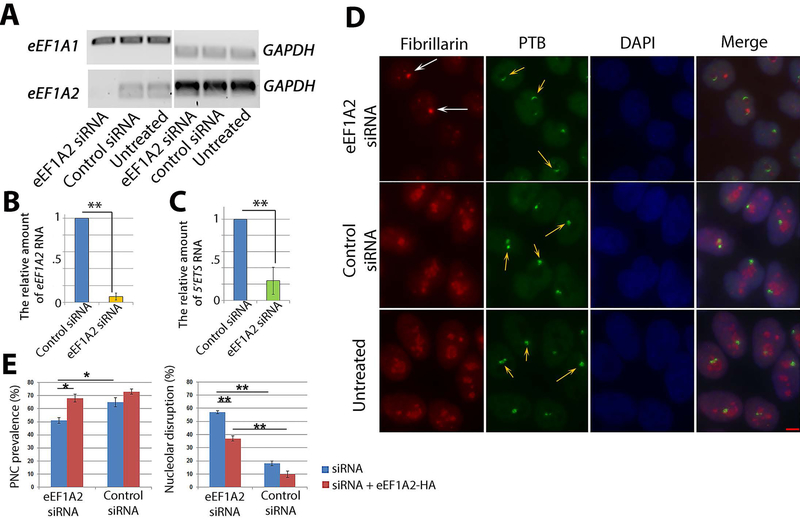
Metastasis remains a leading cause of cancer mortality due to the lack of specific inhibitors against this complex process. To identify compounds selectively targeting the metastatic state, we used the perinucleolar compartment (PNC), a complex nuclear structure associated with metastatic behaviors of cancer cells, as a phenotypic marker for a high-content screen of over 140,000 structurally diverse compounds. Metarrestin, obtained through optimization of a screening hit, disassembles PNCs in multiple cancer cell lines, inhibits invasion in vitro, suppresses metastatic development in three mouse models of human cancer, and extends survival of mice in a metastatic pancreatic cancer xenograft model with no organ toxicity or discernable adverse effects. Metarrestin disrupts the nucleolar structure and inhibits RNA polymerase (Pol) I transcription, at least in part by interacting with the translation elongation factor eEF1A2. Thus, metarrestin represents a potential therapeutic approach for the treatment of metastatic cancer.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
References
-
- Jiang WG, Sanders AJ, Katoh M, Ungefroren H, Gieseler F, Prince M, Thompson SK, Zollo M, Spano D, Dhawan P, Sliva D, Subbarayan PR, Sarkar M, Honoki K, Fujii H, Georgakilas AG, Amedei A, Niccolai E, Amin A, Ashraf SS, Ye L, Helferich WG, Yang X, Boosani CS, Guha G, Ciriolo MR, Aquilano K, Chen S, Azmi AS, Keith WN, Bilsland A, Bhakta D, Halicka D, Nowsheen S, Pantano F, Santini D, Tissue invasion and metastasis: Molecular, biological and clinical perspectives. Semin Cancer Biol 35 Suppl, S244–75 (2015). - PubMed
-
- Weber GF, Why does cancer therapy lack effective anti-metastasis drugs? Cancer Lett 328, 207–11 (2013). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
