

Tumor-associated calreticulin variants functionally compromise the peptide loading complex and impair its recruitment of MHC-I
- PMID: 29769311
- PMCID: PMC6016473
- DOI: 10.1074/jbc.RA118.002836
Tumor-associated calreticulin variants functionally compromise the peptide loading complex and impair its recruitment of MHC-I
Abstract
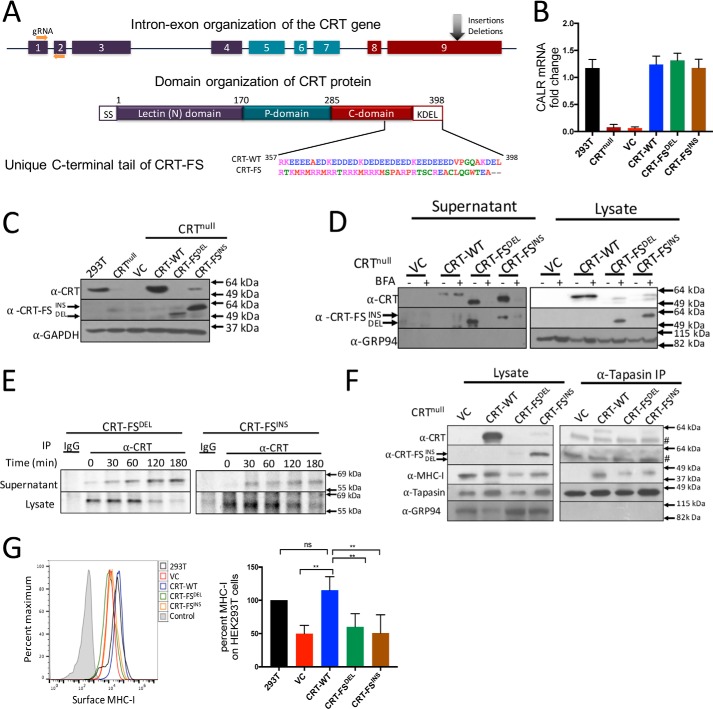
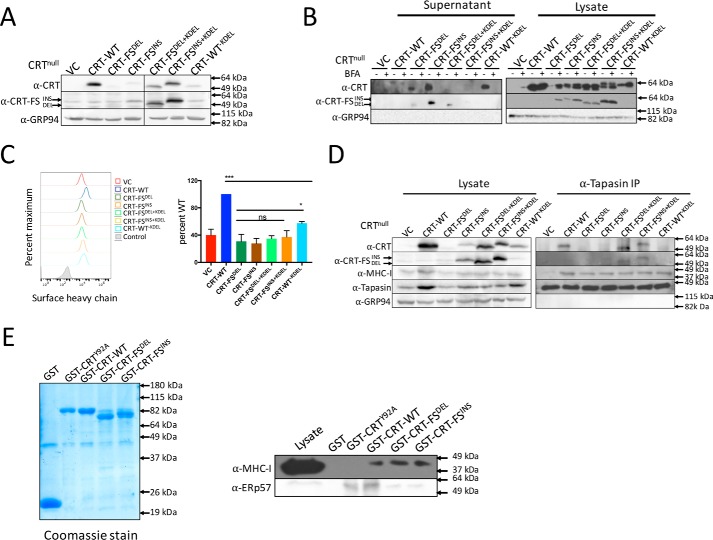
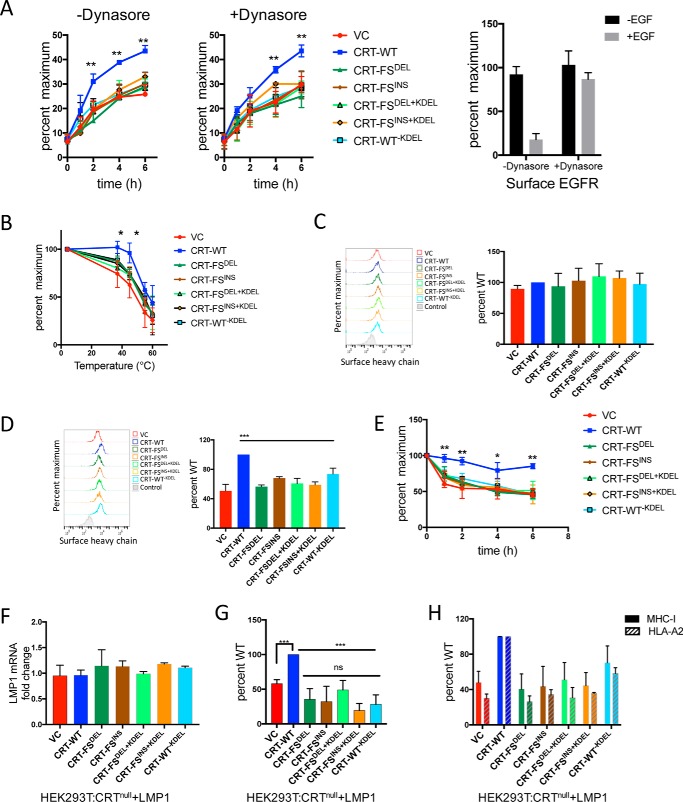
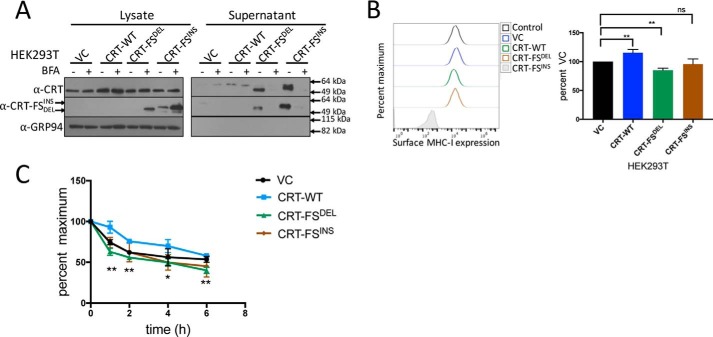
Major histocompatibility complex-I-β2m dimers (MHC-I) bind peptides derived from intracellular proteins, enabling the immune system to distinguish between normal cells and those expressing pathogen-derived or mutant proteins. The peptides bind to MHC-I in the endoplasmic reticulum (ER), and this binding is facilitated by the
Keywords: antigen presentation; antigen processing; calreticulin; major histocompatibility complex (MHC); myeloproliferative neoplasms; peptide-loading complex; protein export; protein secretion; tumor immunology/immunotherapy.
© 2018 Arshad and Cresswell.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
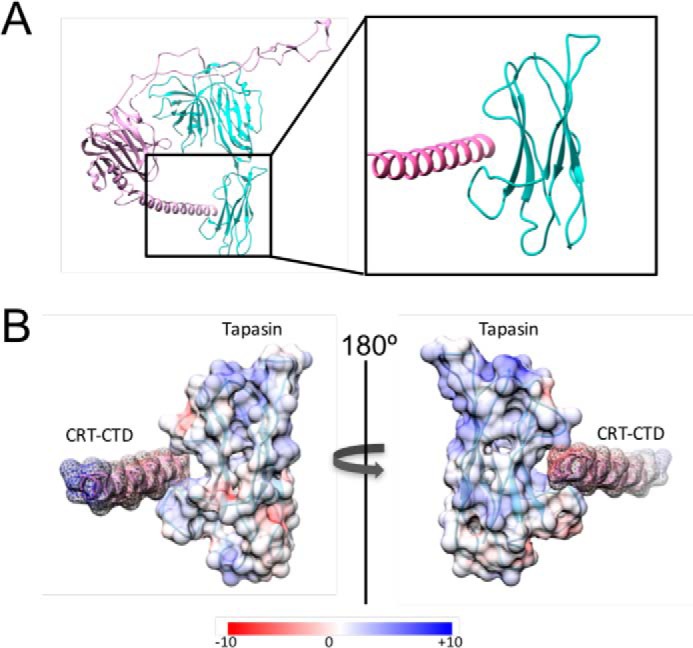
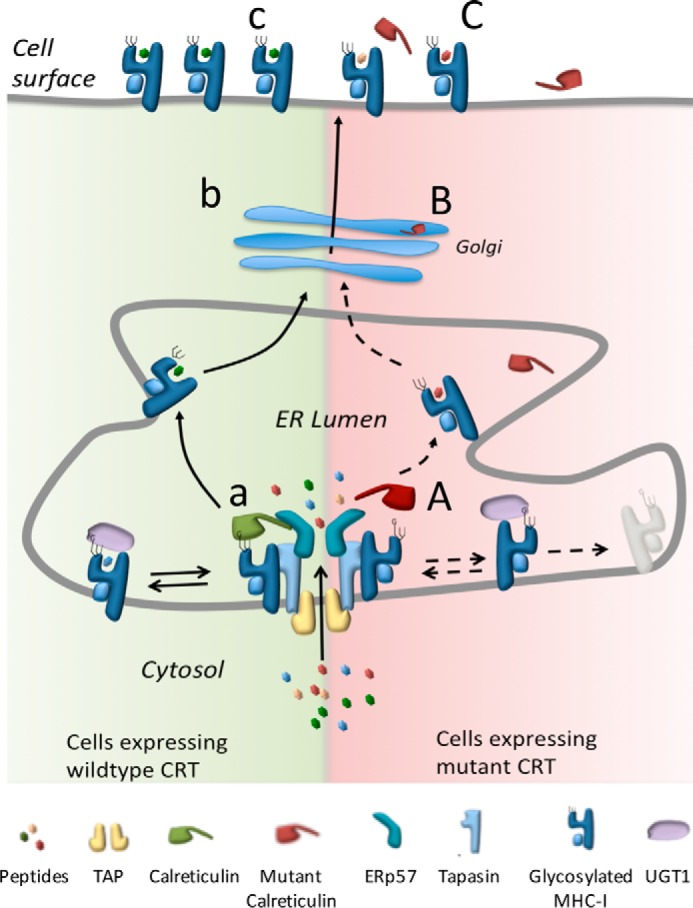
References
-
- Barkal A. A., Weiskopf K., Kao K. S., Gordon S. R., Rosental B., Yiu Y. Y., George B. M., Markovic M., Ring N. G., Tsai J. M., McKenna K. M., Ho P. Y., Cheng R. Z., Chen J. Y., Barkal L. J., Ring A. M., Weissman I. L., and Maute R. L. (2018) Engagement of MHC class I by the inhibitory receptor LILRB1 suppresses macrophages and is a target of cancer immunotherapy. Nat. Immunol. 19, 76–84 10.1038/s41590-017-0004-z - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
