

TLR-adjuvanted nanoparticle vaccines differentially influence the quality and longevity of responses to malaria antigen Pfs25
- PMID: 29769448
- PMCID: PMC6012510
- DOI: 10.1172/jci.insight.120692
TLR-adjuvanted nanoparticle vaccines differentially influence the quality and longevity of responses to malaria antigen Pfs25
Abstract
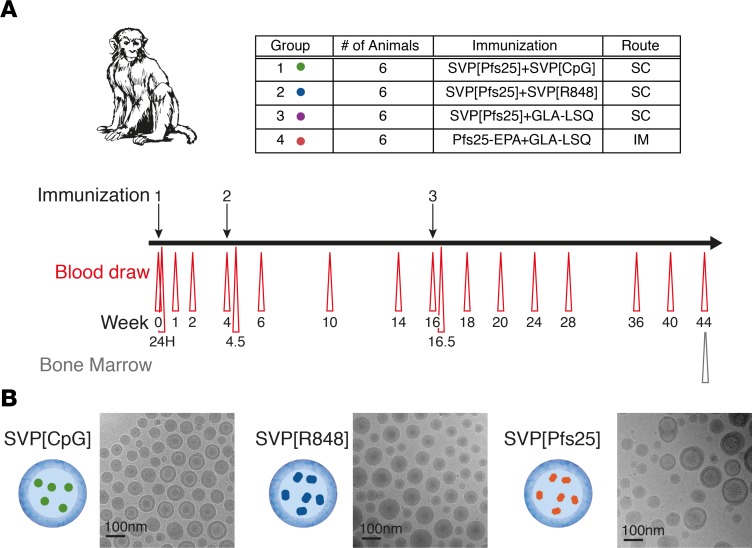
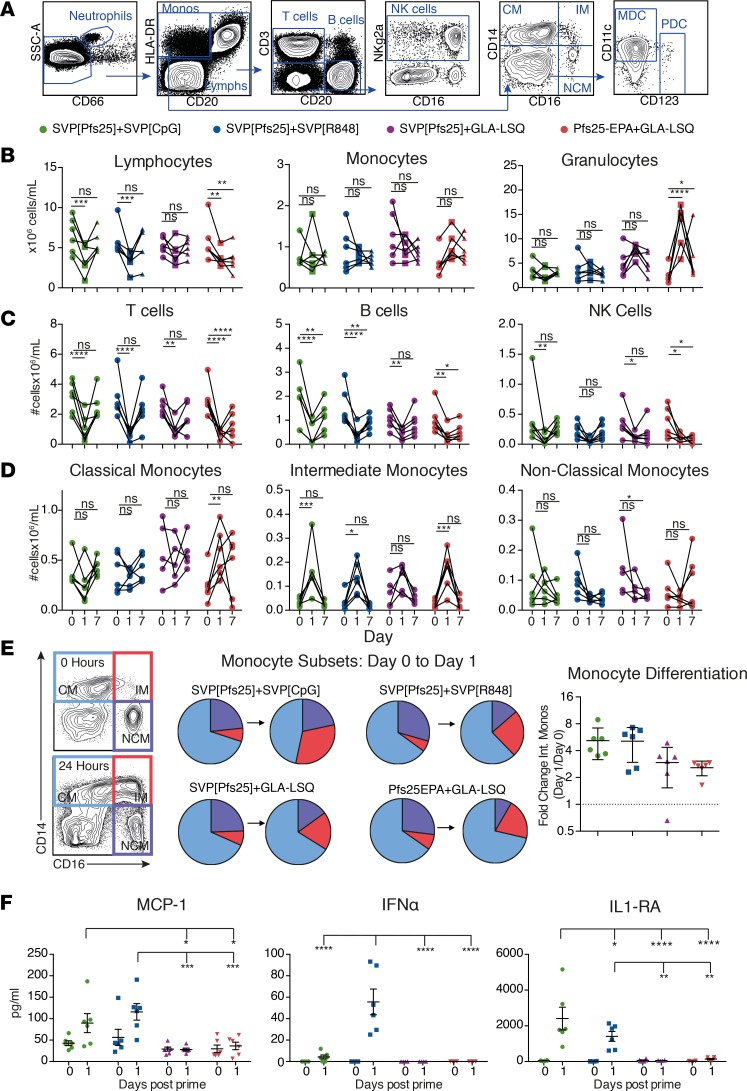
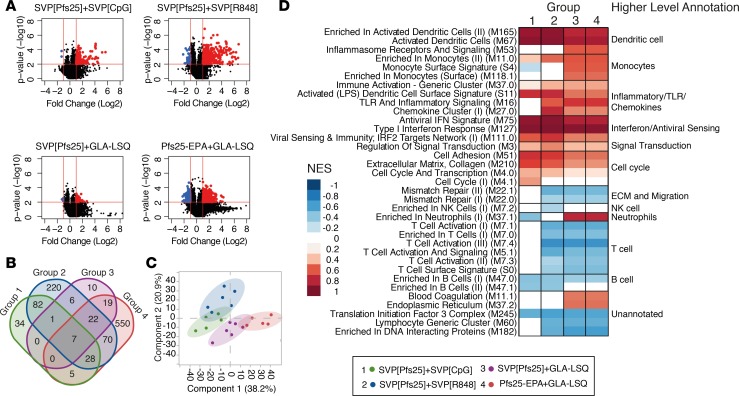
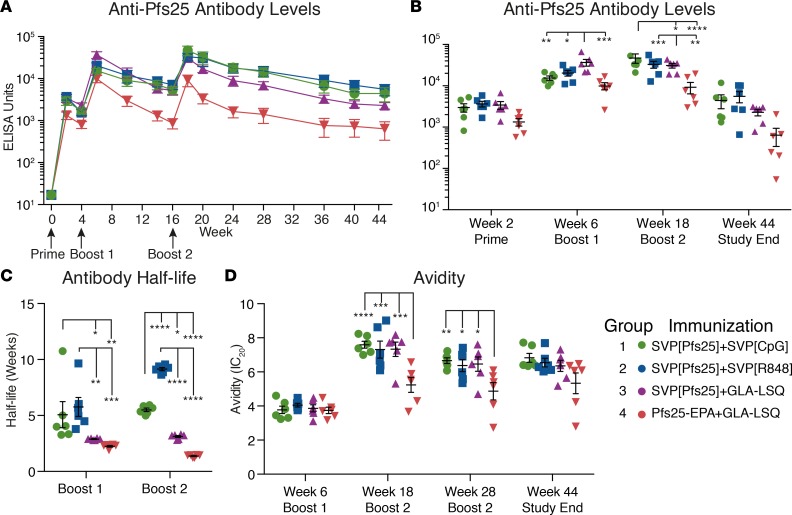
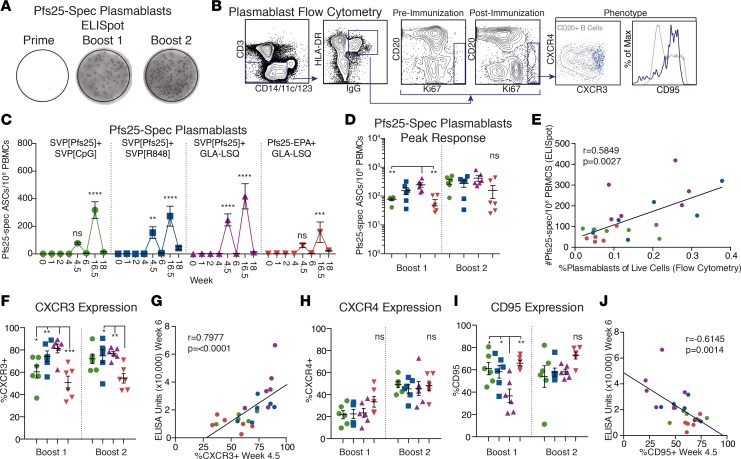
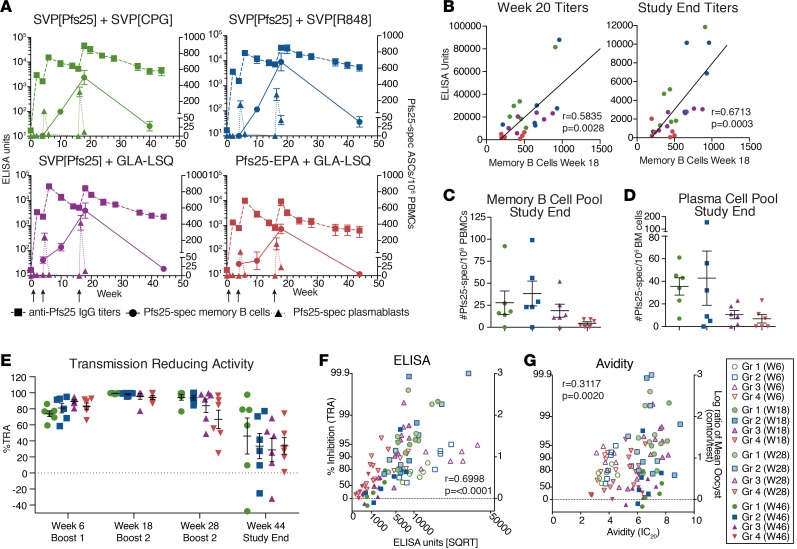
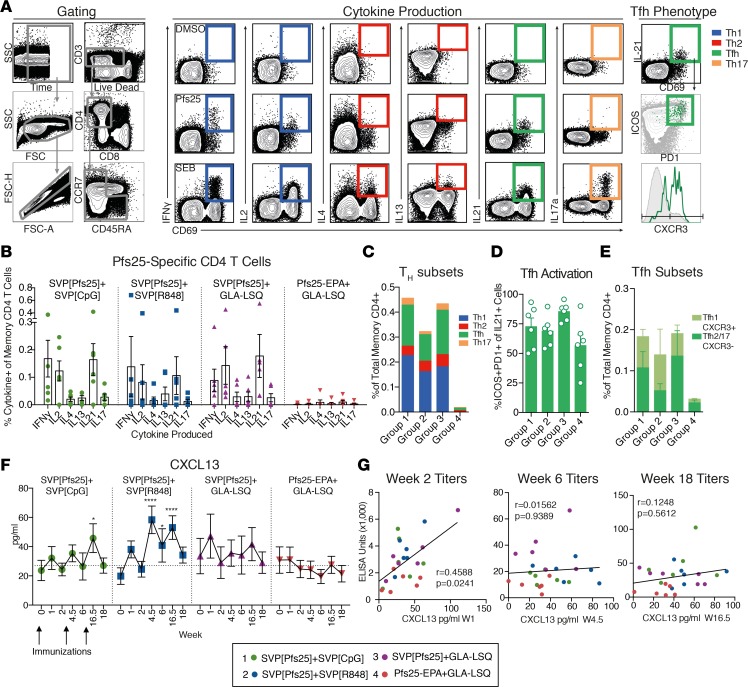
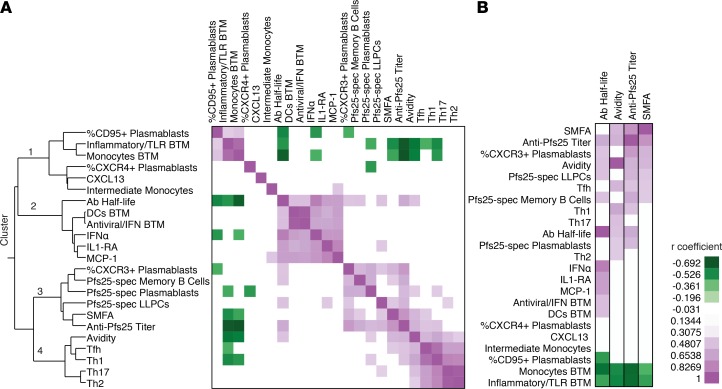
Transmission-blocking vaccines (TBVs) are considered an integral element of malaria eradication efforts. Despite promising evaluations of Plasmodium falciparum Pfs25-based TBVs in mice, clinical trials have failed to induce robust and long-lived Ab titers, in part due to the poorly immunogenic nature of Pfs25. Using nonhuman primates, we demonstrate that multiple aspects of Pfs25 immunity were enhanced by antigen encapsulation in poly(lactic-co-glycolic acid)-based [(PLGA)-based] synthetic vaccine particles (SVP[Pfs25]) and potent TLR-based adjuvants. SVP[Pfs25] increased Ab titers, Pfs25-specific plasmablasts, circulating memory B cells, and plasma cells in the bone marrow when benchmarked against the clinically tested multimeric form Pfs25-EPA given with GLA-LSQ. SVP[Pfs25] also induced the first reported Pfs25-specific circulating Th1 and Tfh cells to our knowledge. Multivariate correlative analysis indicated several mechanisms for the improved Ab responses. While Pfs25-specific B cells were responsible for increasing Ab titers, T cell responses stimulated increased Ab avidity. The innate immune activation differentially stimulated by the adjuvants revealed a strong correlation between type I IFN polarization, induced by R848 and CpG, and increased Ab half-life and longevity. Collectively, the data identify ways to improve vaccine-induced immunity to poorly immunogenic proteins, both by the choice of antigen and adjuvant formulation, and highlight underlying immunological mechanisms.
Keywords: Adaptive immunity; Immunology; Innate immunity; Malaria; Vaccines.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
