

The maize lilliputian1 (lil1) gene, encoding a brassinosteroid cytochrome P450 C-6 oxidase, is involved in plant growth and drought response
- PMID: 29771294
- PMCID: PMC6070094
- DOI: 10.1093/aob/mcy047
The maize lilliputian1 (lil1) gene, encoding a brassinosteroid cytochrome P450 C-6 oxidase, is involved in plant growth and drought response
Abstract
Background and aims: Brassinosteroids (BRs) are plant hormones involved in many developmental processes as well as in plant-environment interactions. Their role was investigated in this study through the analysis of lilliputian1-1 (lil1-1), a dwarf mutant impaired in BR biosynthesis in maize (Zea mays).
Methods: We isolated lil1-1 through transposon tagging in maize. The action of lil1 was investigated through morphological and genetic analysis. Moreover, by comparing lil1-1 mutant and wild-type individuals grown under drought stress, the effect of BR reduction on the response to drought stress was examined.
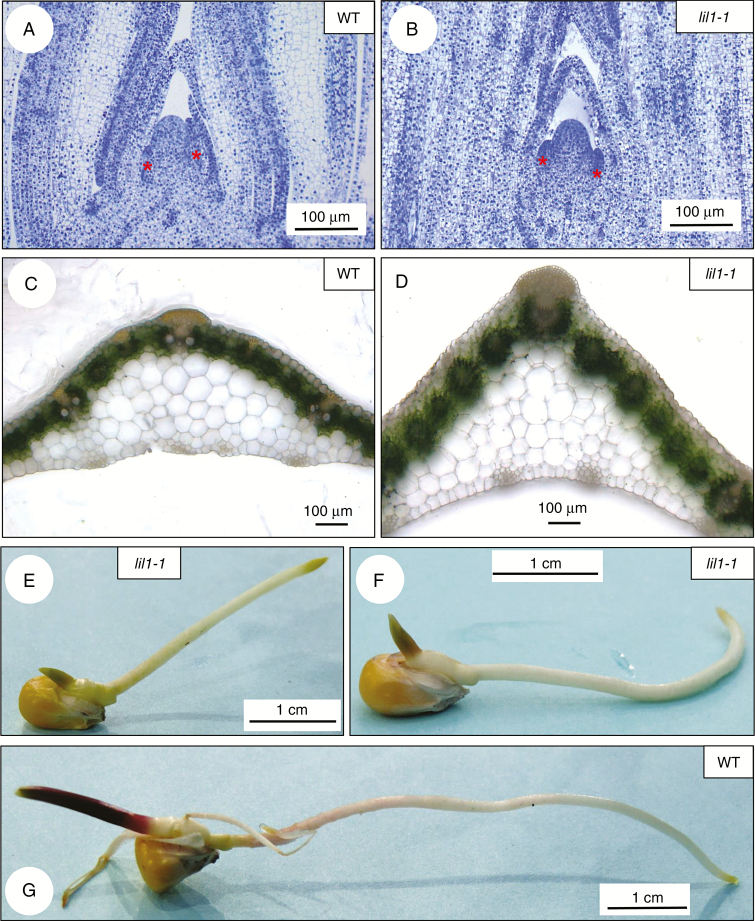
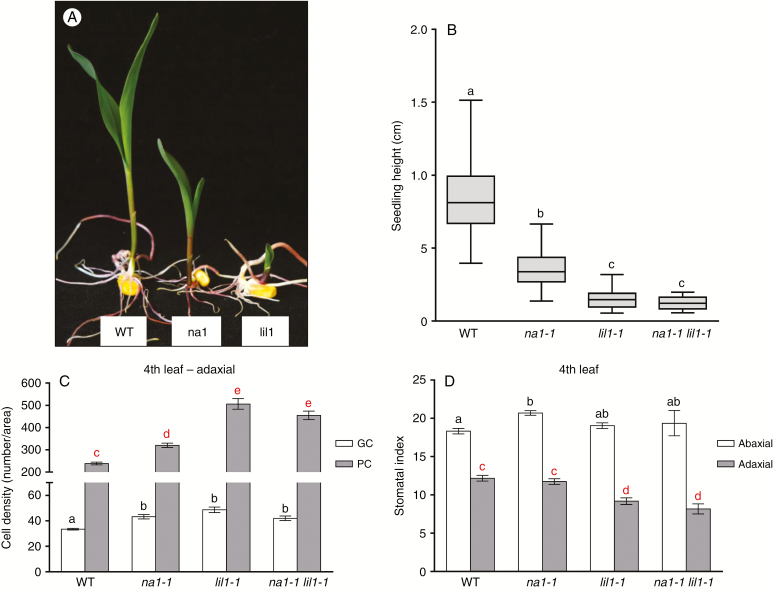
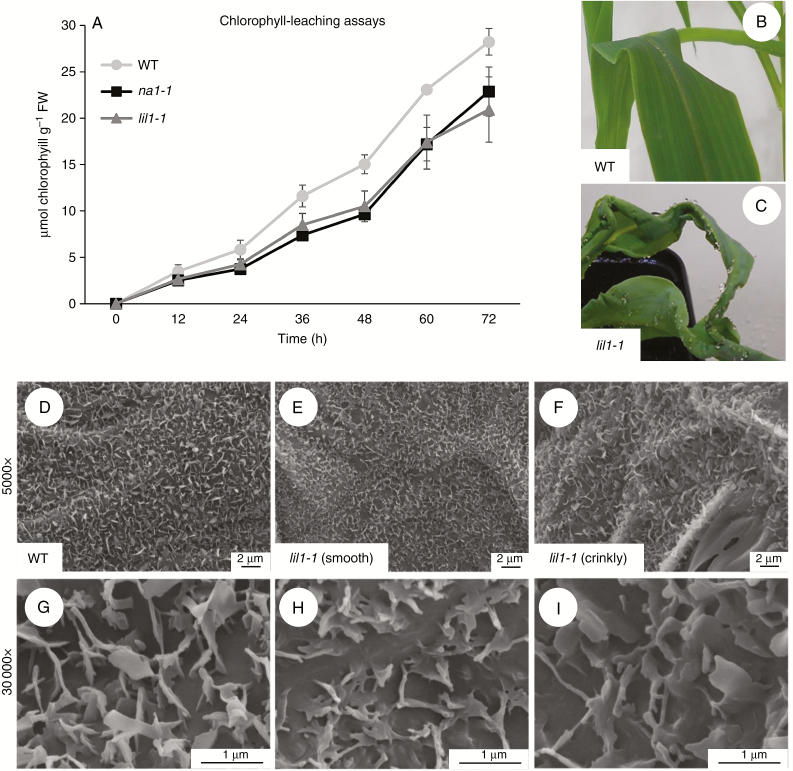
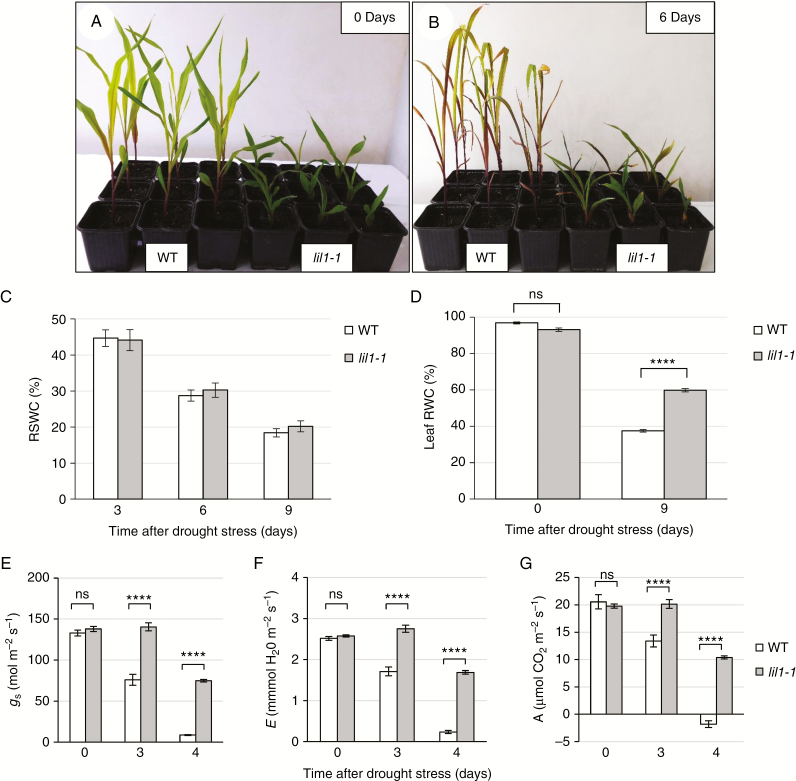
Key results: lil1-1 is a novel allele of the brassinosteroid-deficient dwarf1 (brd1) gene, encoding a brassinosteroid C-6 oxidase. We show in this study that lil1 is epistatic to nana plant1 (na1), a BR gene involved in earlier steps of the pathway. The lill-1 mutation causes alteration in the root gravitropic response, leaf epidermal cell density, epicuticular wax deposition and seedling adaptation to water scarcity conditions.
Conclusions: Lack of active BR molecules in maize causes a pleiotropic effect on plant development and improves seedling tolerance of drought. BR-deficient maize mutants can thus be instrumental in unravelling novel mechanisms on which plant adaptations to abiotic stress are based.
Figures

References
-
- Amzallag GN, Vaisman J. 2006. Influence of brassinosteroids on initiation of the root gravitropic response in Pisum sativum seedlings. Biologia Plantarum 50: 283–286.
-
- Bajguz A, Hayat S. 2009. Effects of brassinosteroids on the plant responses to environmental stresses. Plant Physiology and Biochemistry 47: 1–8. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
