

Elevated temperature increases meiotic crossover frequency via the interfering (Type I) pathway in Arabidopsis thaliana
- PMID: 29771908
- PMCID: PMC5976207
- DOI: 10.1371/journal.pgen.1007384
Elevated temperature increases meiotic crossover frequency via the interfering (Type I) pathway in Arabidopsis thaliana
Abstract
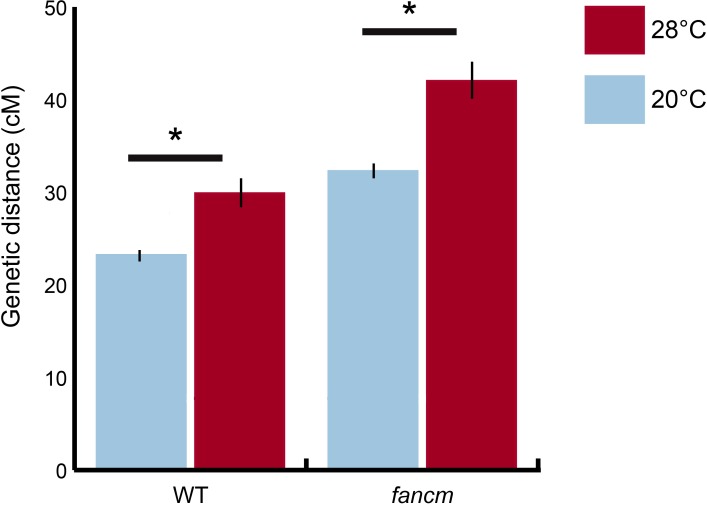
For most eukaryotes, sexual reproduction is a fundamental process that requires meiosis. In turn, meiosis typically depends on a reciprocal exchange of DNA between each pair of homologous chromosomes, known as a crossover (CO), to ensure proper chromosome segregation. The frequency and distribution of COs are regulated by intrinsic and extrinsic environmental factors, but much more is known about the molecular mechanisms governing the former compared to the latter. Here we show that elevated temperature induces meiotic hyper-recombination in Arabidopsis thaliana and we use genetic analysis with mutants in different recombination pathways to demonstrate that the extra COs are derived from the major Type I interference sensitive pathway. We also show that heat-induced COs are not the result of an increase in DNA double-strand breaks and that the hyper-recombinant phenotype is likely specific to thermal stress rather than a more generalized stress response. Taken together, these findings provide initial mechanistic insight into how environmental cues modulate plant meiotic recombination and may also offer practical applications.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
-
- Mercier R, Mézard C, Jenczewski E, Macaisne N, Grelon M. The Molecular Biology of Meiosis in Plants. Annu Rev Plant Biol. 2015; 66(1):297–327. doi: 10.1146/annurev-arplant-050213-035923 - DOI - PubMed
-
- Keeney S, Giroux CN, Kleckner N. Meiosis-specific DNA double-strand breaks are catalyzed by Spo11, a member of a widely conserved protein family. Cell. 1997; 88(3):375–84. doi: 10.1016/S0092-8674(00)81876-0 - DOI - PubMed
-
- Copenhaver GP, Browne WE, Preuss D. Assaying genome-wide recombination and centromere functions with Arabidopsis tetrads. Proc Natl Acad Sci U S A. 1998; 95 (1) 247–52. doi: 10.1073/pnas.95.1.247 - DOI - PMC - PubMed
-
- Mercier R, Jolivet S, Vezon D, Huppe E, Chelysheva L, Giovanni M, et al. Two meiotic crossover classes cohabit in Arabidopsis: one is dependent on MER3, whereas the other one is not. Curr Biol. 2005; 15(8):692–701. doi: 10.1016/j.cub.2005.02.056 - DOI - PubMed
-
- Chelysheva L, Gendrot G, Vezon D, Doutriaux M-P, Mercier R, Grelon M. Zip4/Spo22 is required for class I CO formation but not for synapsis completion in Arabidopsis thaliana. PLoS Genet. 2007; 3(5):e83 doi: 10.1371/journal.pgen.0030083 - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
