

Induction of influenza-specific local CD8 T-cells in the respiratory tract after aerosol delivery of vaccine antigen or virus in the Babraham inbred pig
- PMID: 29772011
- PMCID: PMC5957346
- DOI: 10.1371/journal.ppat.1007017
Induction of influenza-specific local CD8 T-cells in the respiratory tract after aerosol delivery of vaccine antigen or virus in the Babraham inbred pig
Abstract
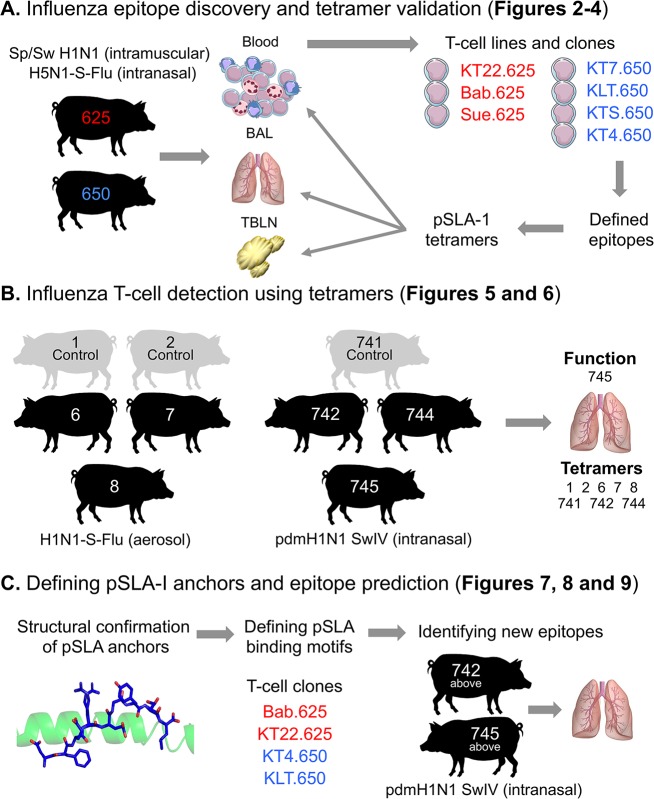
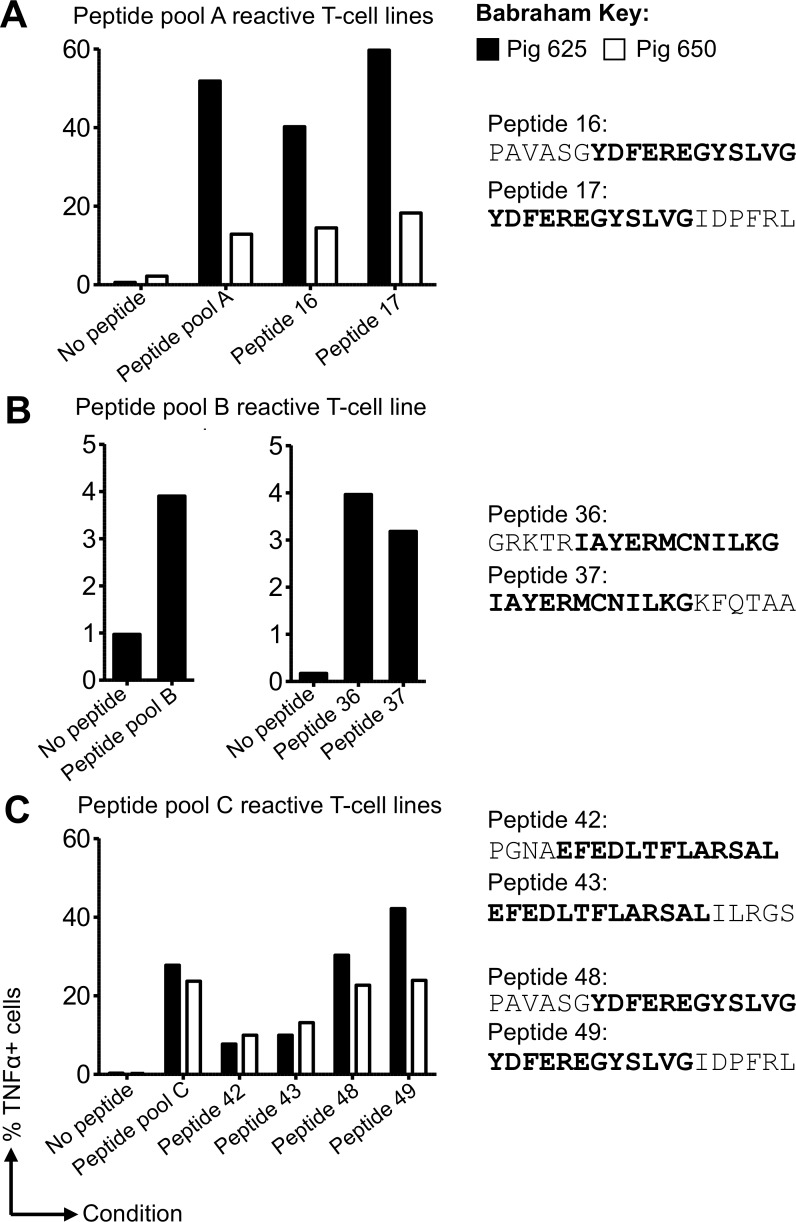
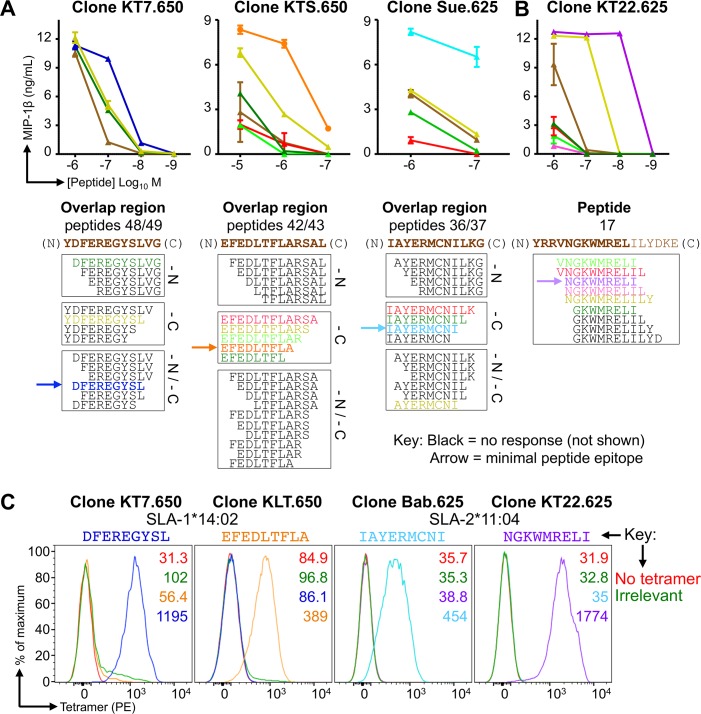
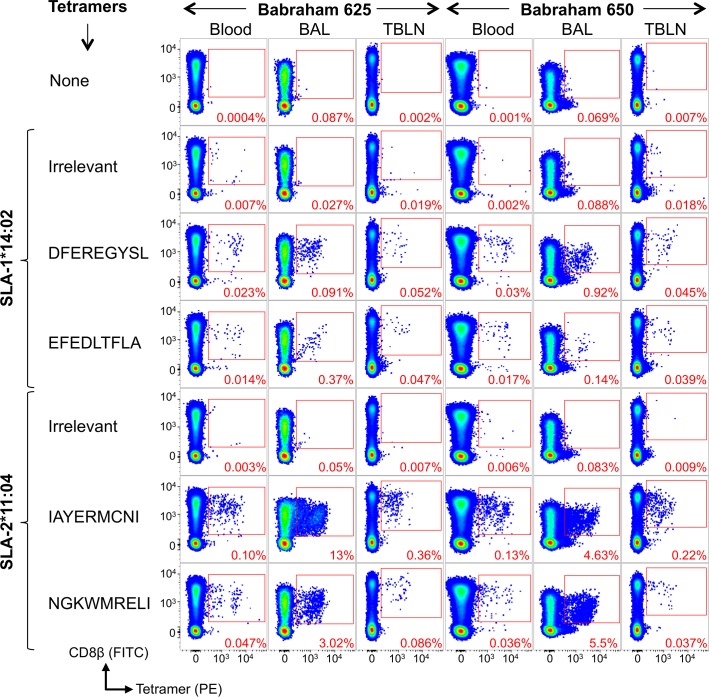
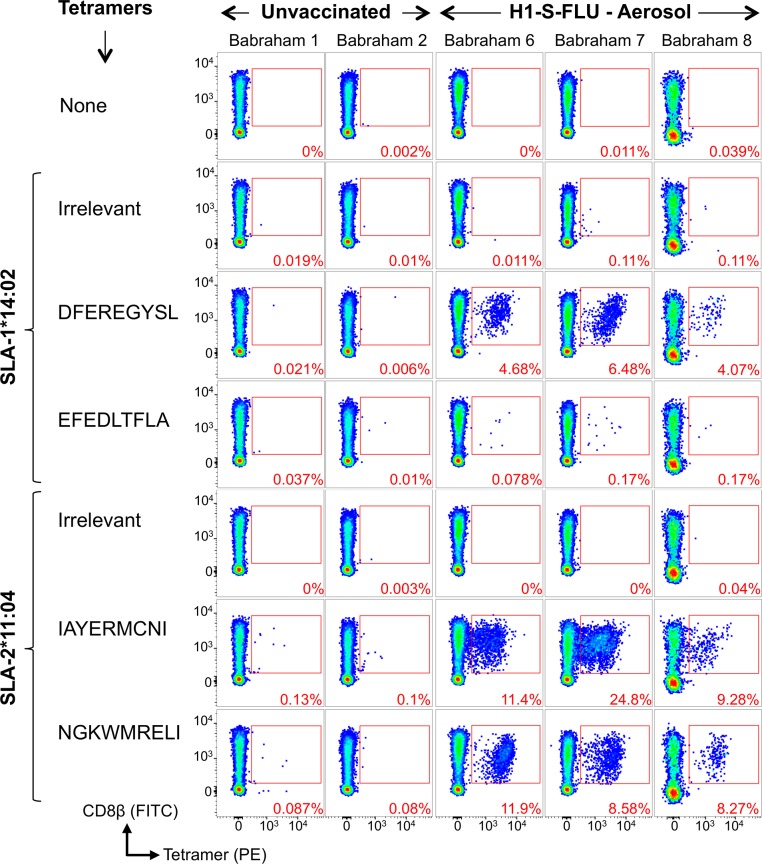
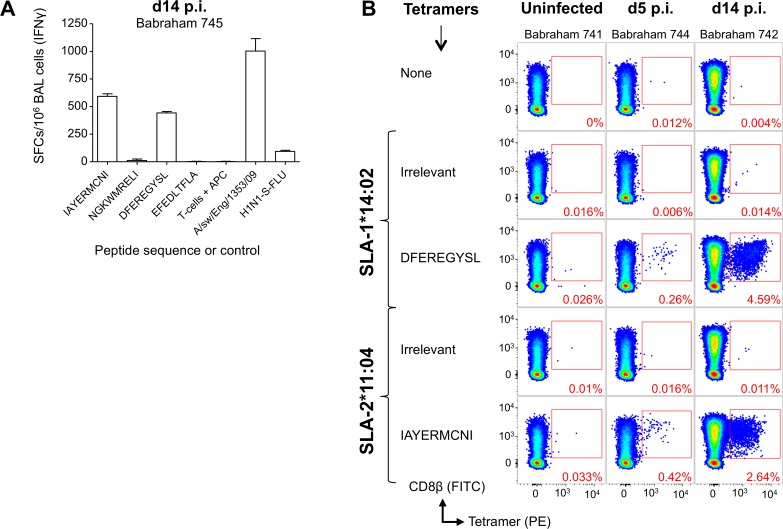
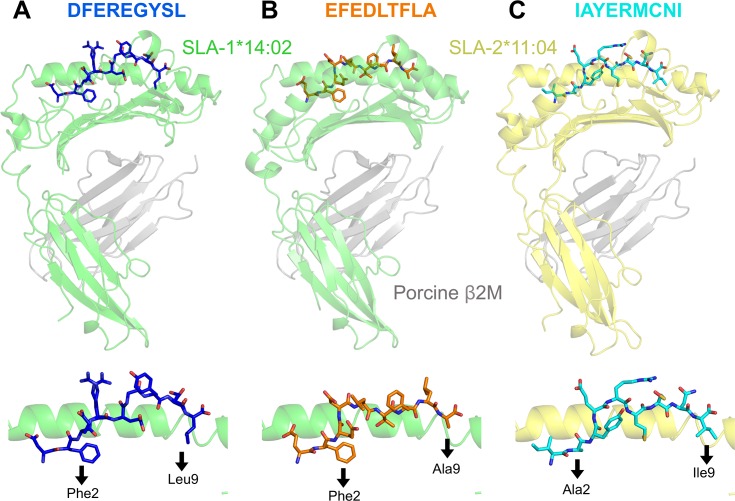
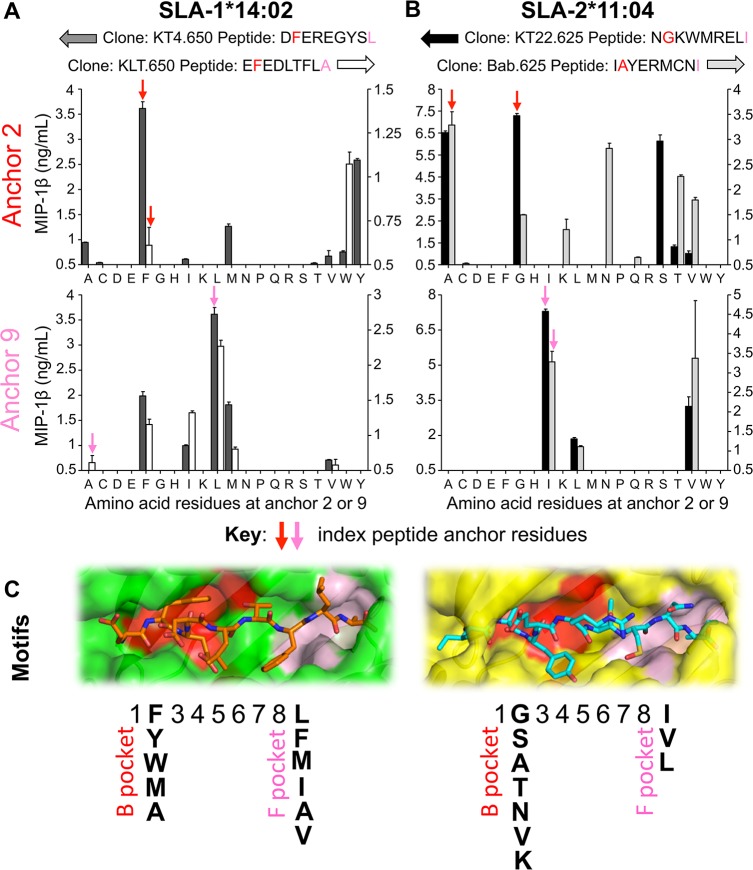
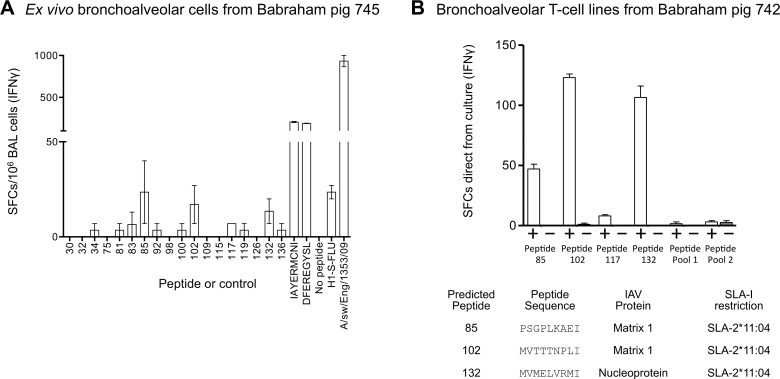
There is increasing evidence that induction of local immune responses is a key component of effective vaccines. For respiratory pathogens, for example tuberculosis and influenza, aerosol delivery is being actively explored as a method to administer vaccine antigens. Current animal models used to study respiratory pathogens suffer from anatomical disparity with humans. The pig is a natural and important host of influenza viruses and is physiologically more comparable to humans than other animal models in terms of size, respiratory tract biology and volume. It may also be an important vector in the birds to human infection cycle. A major drawback of the current pig model is the inability to analyze antigen-specific CD8+ T-cell responses, which are critical to respiratory immunity. Here we address this knowledge gap using an established in-bred pig model with a high degree of genetic identity between individuals, including the MHC (Swine Leukocyte Antigen (SLA)) locus. We developed a toolset that included long-term in vitro pig T-cell culture and cloning and identification of novel immunodominant influenza-derived T-cell epitopes. We also generated structures of the two SLA class I molecules found in these animals presenting the immunodominant epitopes. These structures allowed definition of the primary anchor points for epitopes in the SLA binding groove and established SLA binding motifs that were used to successfully predict other influenza-derived peptide sequences capable of stimulating T-cells. Peptide-SLA tetramers were constructed and used to track influenza-specific T-cells ex vivo in blood, the lungs and draining lymph nodes. Aerosol immunization with attenuated single cycle influenza viruses (S-FLU) induced large numbers of CD8+ T-cells specific for conserved NP peptides in the respiratory tract. Collectively, these data substantially increase the utility of pigs as an effective model for studying protective local cellular immunity against respiratory pathogens.
Conflict of interest statement
AMT is named on a European patent (publication no. EP2758525 A2, published July 30, 2014) concerning the use of S-FLU as a vaccine. AKS. is an inventor of patent WO 2010032022, “Use of protein kinase inhibitor to detect immune cells, such as T-cells. The other authors have no financial conflicts of interest.
Figures
References
-
- Molinari N-AM, Ortega-Sanchez IR, Messonnier ML, Thompson WW, Wortley PM, Weintraub E, et al. The annual impact of seasonal influenza in the US: Measuring disease burden and costs. Vaccine. 2007;25: 5086–5096. doi: 10.1016/j.vaccine.2007.03.046 - DOI - PubMed
-
- Tchilian E, Holzer B. Harnessing Local Immunity for an Effective Universal Swine Influenza Vaccine. Viruses. Multidisciplinary Digital Publishing Institute; 2017;9: 98 doi: 10.3390/v9050098 - DOI - PMC - PubMed
-
- Paules CI, Marston HD, Eisinger RW, Baltimore D, Fauci AS. The Pathway to a Universal Influenza Vaccine. Immunity. 2017. pp. 599–603. doi: 10.1016/j.immuni.2017.09.007 - DOI - PubMed
-
- McMichael AJ, Gotch FM, Noble GR, Beare PAS. Cytotoxic T-Cell Immunity to Influenza. N Engl J Med. 1983;309: 13–17. doi: 10.1056/NEJM198307073090103 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
