

Perlecan, a heparan sulfate proteoglycan, regulates systemic metabolism with dynamic changes in adipose tissue and skeletal muscle
- PMID: 29773865
- PMCID: PMC5958100
- DOI: 10.1038/s41598-018-25635-x
Perlecan, a heparan sulfate proteoglycan, regulates systemic metabolism with dynamic changes in adipose tissue and skeletal muscle
Abstract
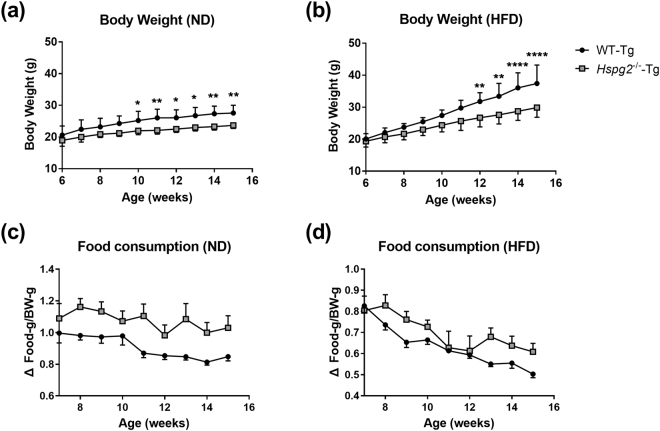
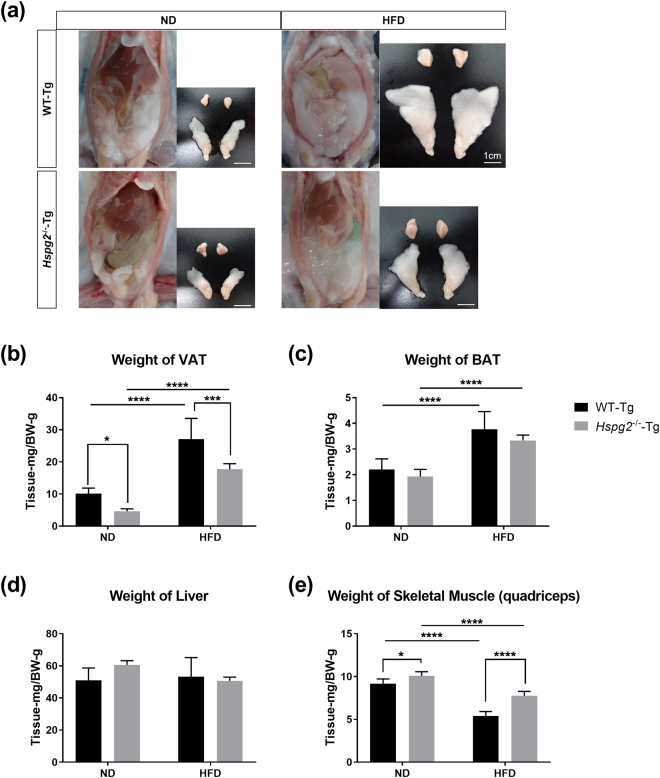
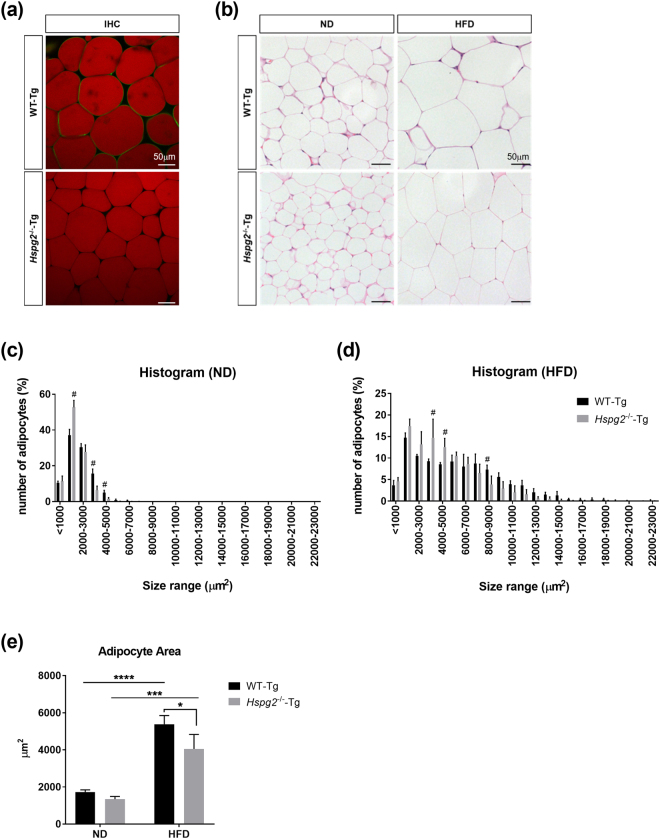
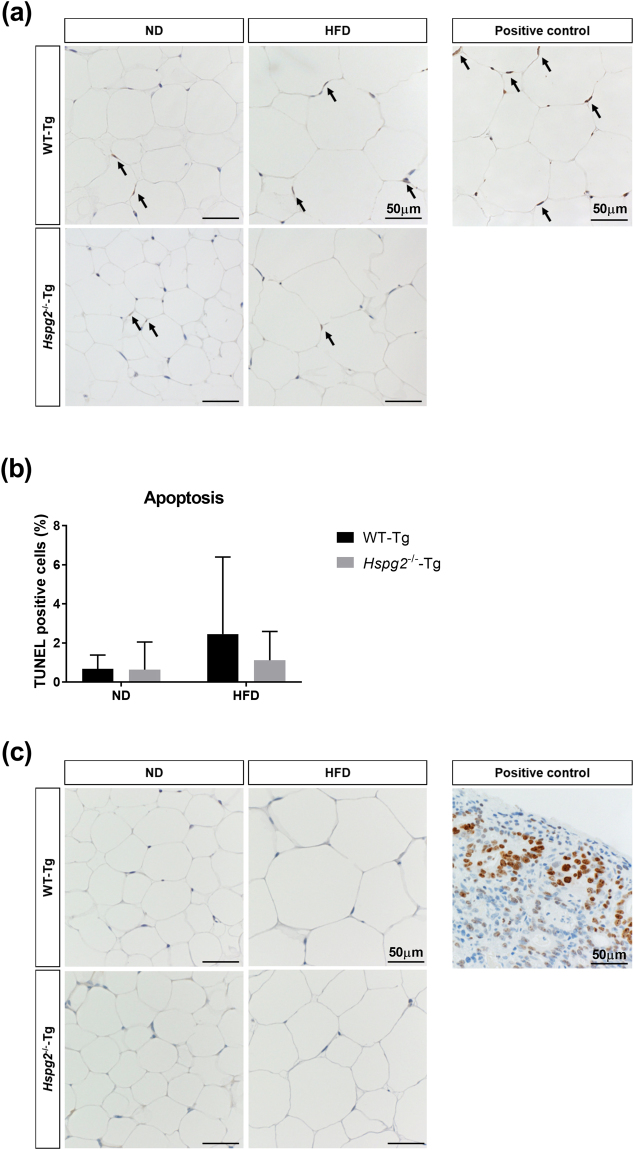
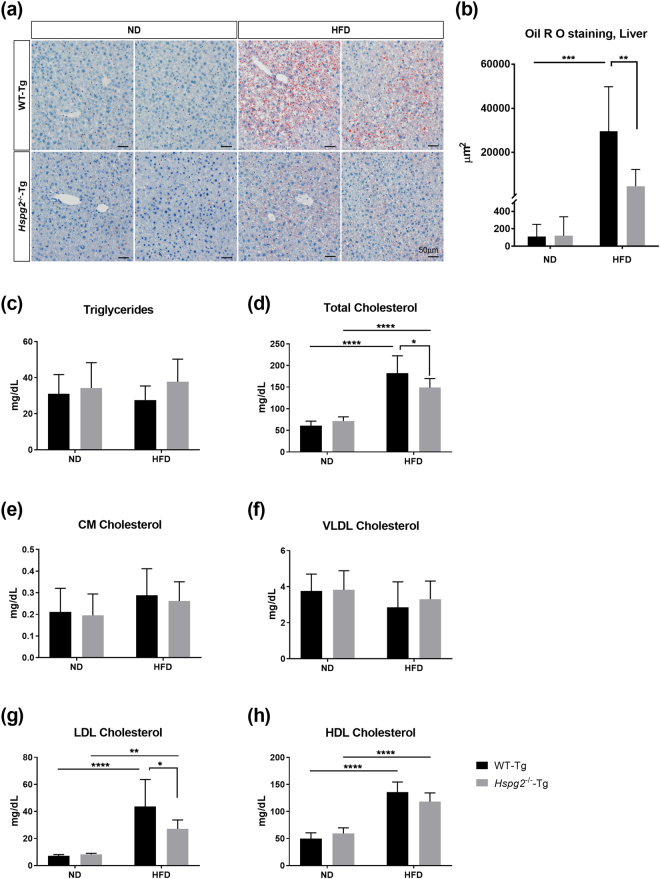
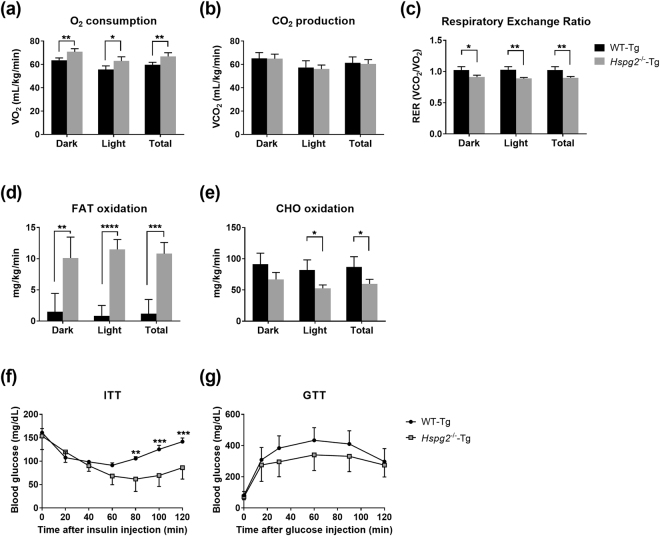
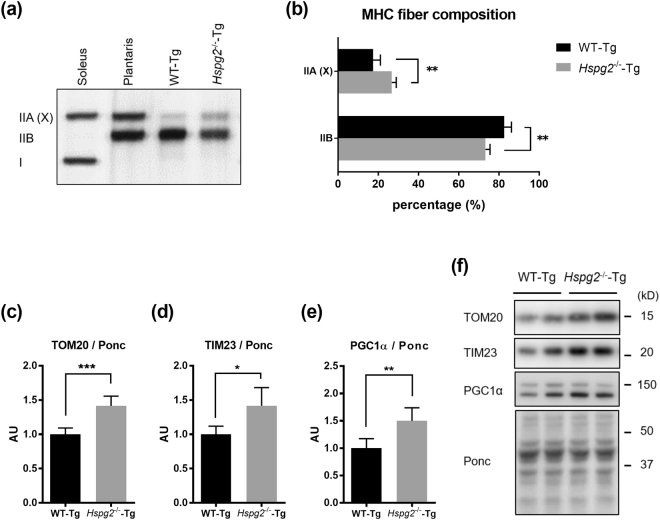
Perlecan (HSPG2), a heparan sulfate proteoglycan, is a component of basement membranes and participates in a variety of biological activities. Here, we show physiological roles of perlecan in both obesity and the onset of metabolic syndrome. The perinatal lethality-rescued perlecan knockout (Hspg2-/--Tg) mice showed a smaller mass and cell size of white adipose tissues than control (WT-Tg) mice. Abnormal lipid deposition, such as fatty liver, was not detected in the Hspg2-/--Tg mice, and those mice also consumed more fat as an energy source, likely due to their activated fatty acid oxidation. In addition, the Hspg2-/--Tg mice demonstrated increased insulin sensitivity. Molecular analysis revealed the significantly relatively increased amount of the muscle fiber type IIA (X) isoform and a larger quantity of mitochondria in the skeletal muscle of Hspg2-/--Tg mice. Furthermore, the perlecan-deficient skeletal muscle also had elevated levels of peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC1α) protein. PGC1α expression is activated by exercise, and induces mitochondrial biosynthesis. Thus, perlecan may act as a mechano-regulator of catabolism of both lipids and glucose by shifting the muscle fiber composition to oxidative fibers. Our data suggest that downregulation of perlecan is a promising strategy to control metabolic syndrome.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
