

Enhanced Sampling of Interdomain Motion Using Map-Restrained Langevin Dynamics and NMR: Application to Pin1
- PMID: 29775635
- PMCID: PMC6002765
- DOI: 10.1016/j.jmb.2018.05.007
Enhanced Sampling of Interdomain Motion Using Map-Restrained Langevin Dynamics and NMR: Application to Pin1
Abstract
Many signaling proteins consist of globular domains connected by flexible linkers that allow for substantial domain motion. Because these domains often serve as complementary functional modules, the possibility of functionally important domain motions arises. To explore this possibility, we require knowledge of the ensemble of protein conformations sampled by interdomain motion. Measurements of NMR residual dipolar couplings (RDCs) of backbone HN bonds offer a per-residue characterization of interdomain dynamics, as the couplings are sensitive to domain orientation. A challenge in reaching this potential is the need to interpret the RDCs as averages over dynamic ensembles of domain conformations. Here, we address this challenge by introducing an efficient protocol for generating conformational ensembles appropriate for flexible, multi-domain proteins. The protocol uses map-restrained self-guided Langevin dynamics simulations to promote collective, interdomain motion while restraining the internal domain motion to near rigidity. Critically, the simulations retain an all-atom description for facile inclusion of site-specific NMR RDC restraints. The result is the rapid generation of conformational ensembles consistent with the RDC data. We illustrate this protocol on human Pin1, a two-domain peptidyl-prolyl isomerase relevant for cancer and Alzheimer's disease. The results include the ensemble of domain orientations sampled by Pin1, as well as those of a dysfunctional variant, I28A-Pin1. The differences between the ensembles corroborate our previous spin relaxation results that showed weakened interdomain contact in the I28A variant relative to wild type. Our protocol extends our abilities to explore the functional significance of protein domain motions.
Keywords: domain motion; ensemble description; multi-conformational fitting; residual dipolar couplings; self-guided Langevin dynamics.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Figures

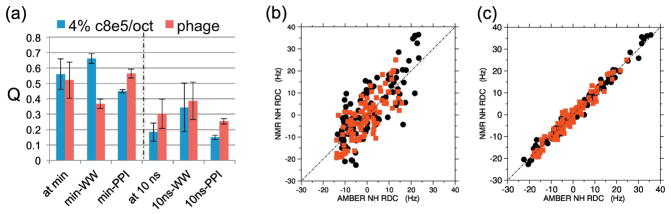


 ) indicate the seven I28A conformers identified by GA/multi-conformational search; stacked diamonds/x’s are populations in nearly the same bins. Panels (G and H) compare conformations sampled by the raw WT-Pin1 versus I28A-Pin1 ensembles (i.e. MapSGLD-NMR simulations producing 90,000 conformations). Yellow-orange ribbons indicate the most frequently sampled conformation; magenta indicates a less populated conformation.
) indicate the seven I28A conformers identified by GA/multi-conformational search; stacked diamonds/x’s are populations in nearly the same bins. Panels (G and H) compare conformations sampled by the raw WT-Pin1 versus I28A-Pin1 ensembles (i.e. MapSGLD-NMR simulations producing 90,000 conformations). Yellow-orange ribbons indicate the most frequently sampled conformation; magenta indicates a less populated conformation.

References
-
- Pufall MA, Graves BJ. Autoinhibitory domains: modular effectors of cellular regulation. Annu Rev Cell Dev Biol. 2002;18:421–62. - PubMed
-
- Bhattacharyya RP, Remenyi A, Yeh BJ, Lim WA. Domains, motifs, and scaffolds: The role of modular interactions in the evolution and wiring of cell signaling circuits. Annual Review of Biochemistry. 2006:655–80. - PubMed
-
- Tolman JR, Flanagan JM, Kennedy MA, Prestegard JH. NMR evidence for slow collective motions in cyanometmyoglobin. Nature Structural Biology. 1997;4:292–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
