

Sequence diversity of tubulin isotypes in regulation of the mitochondrial voltage-dependent anion channel
- PMID: 29777059
- PMCID: PMC6052224
- DOI: 10.1074/jbc.RA117.001569
Sequence diversity of tubulin isotypes in regulation of the mitochondrial voltage-dependent anion channel
Abstract
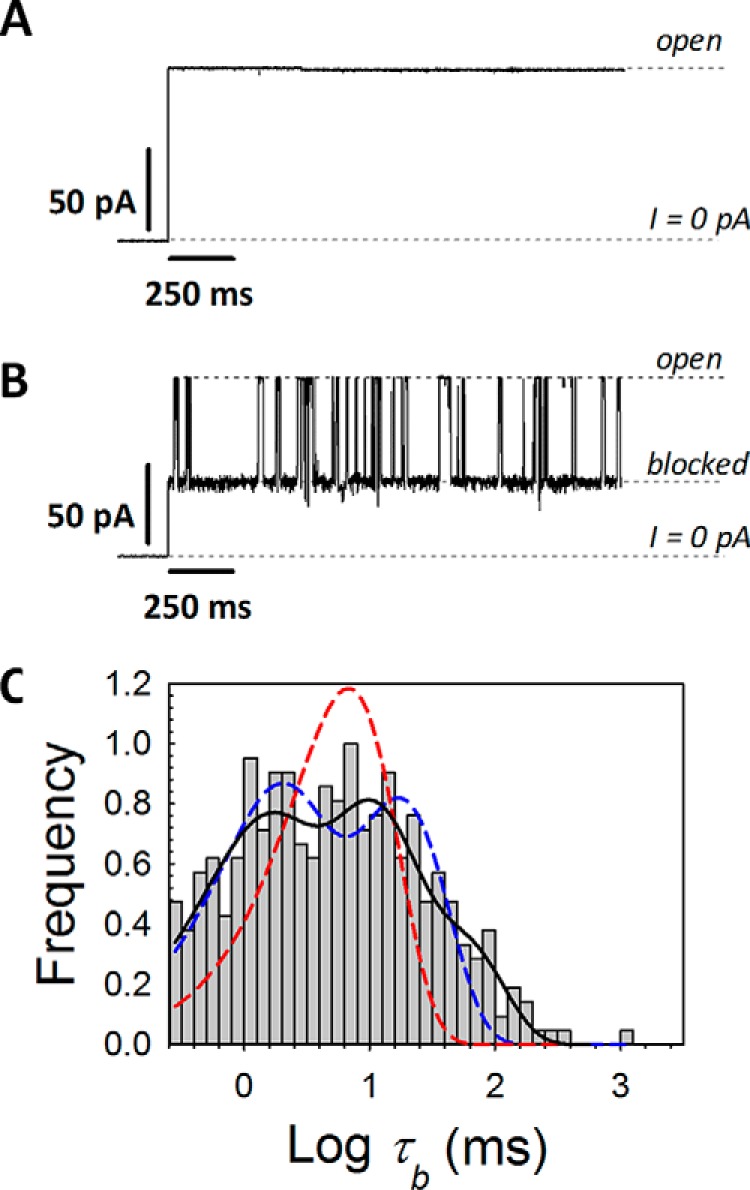
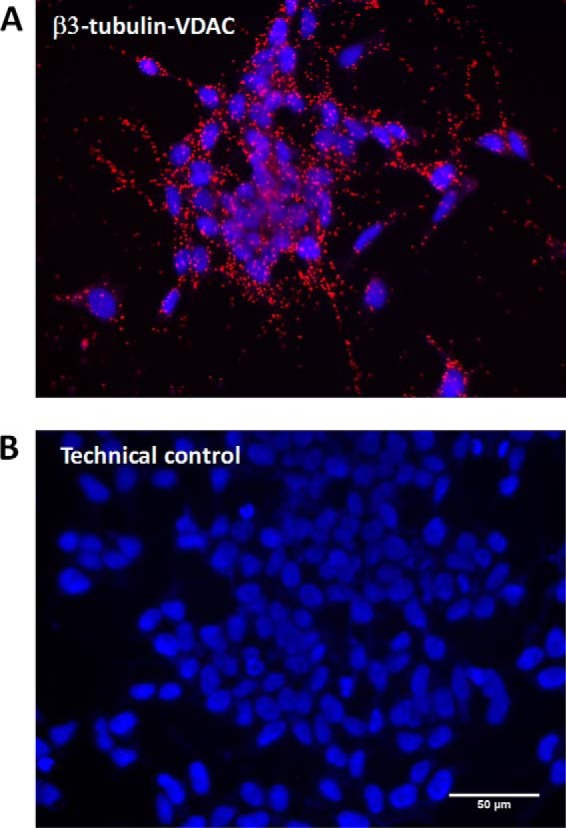
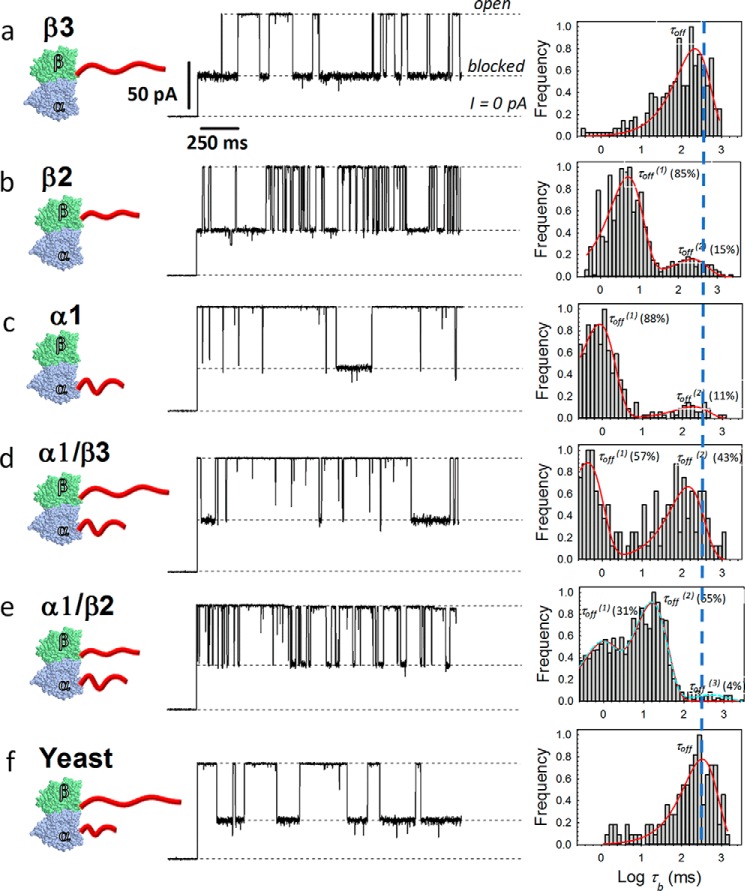
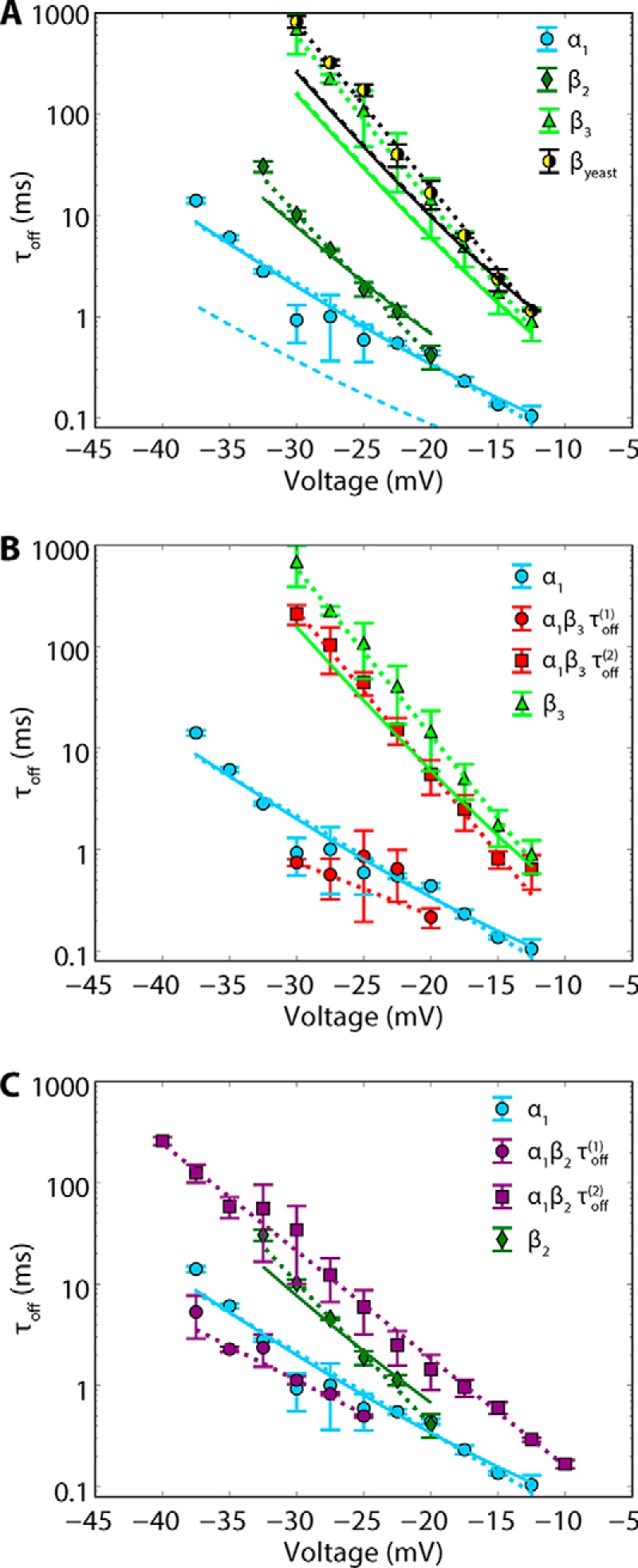
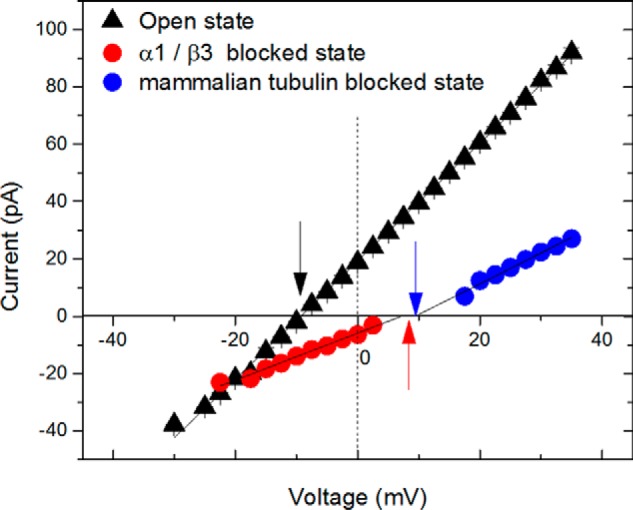
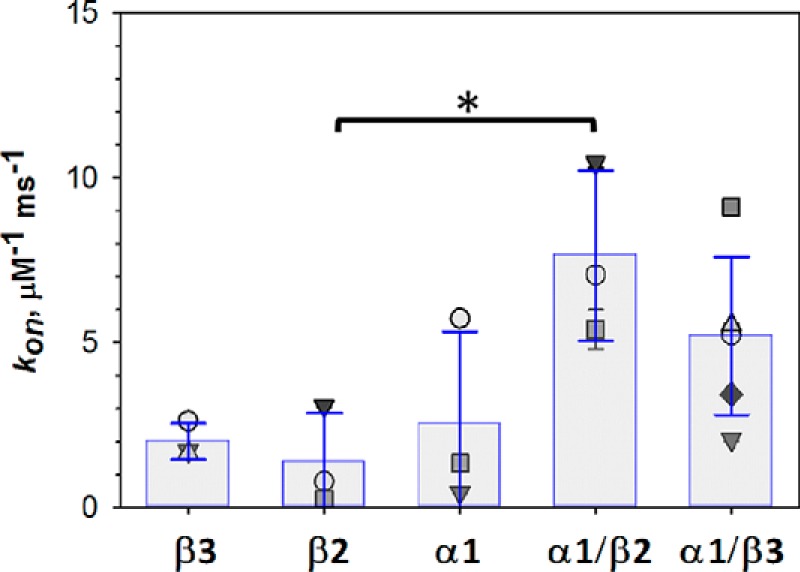
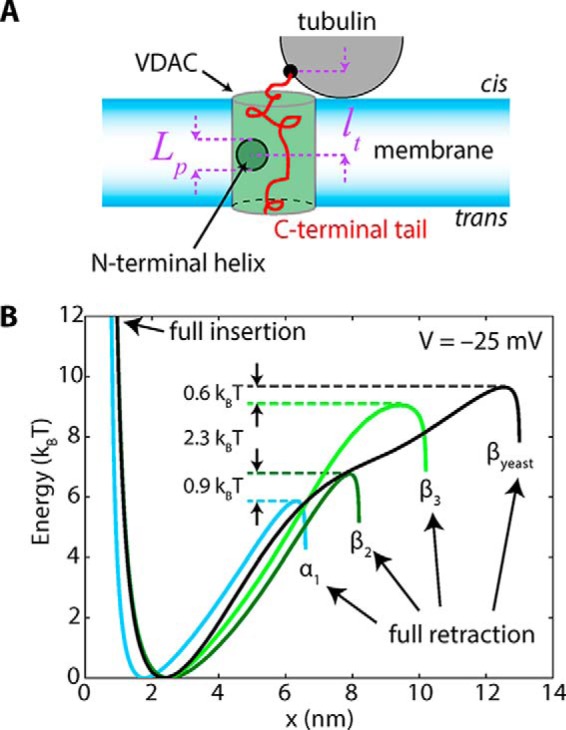
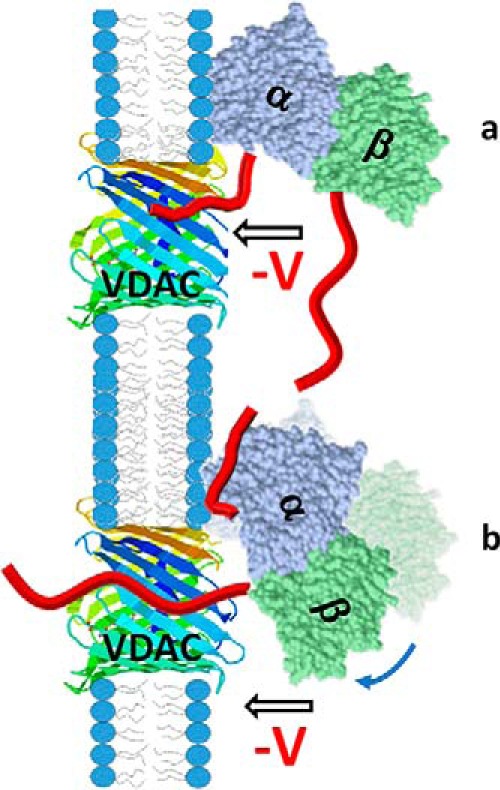
The microtubule protein tubulin is a heterodimer comprising α/β subunits, in which each subunit features multiple isotypes in vertebrates. For example, seven α-tubulin and eight β-tubulin isotypes in the human tubulin gene family vary mostly in the length and primary sequence of the disordered anionic carboxyl-terminal tails (CTTs). The biological reason for such sequence diversity remains a topic of vigorous enquiry. Here, we demonstrate that it may be a key feature of tubulin's role in regulation of the permeability of the mitochondrial outer membrane voltage-dependent anion channel (VDAC). Using recombinant yeast α/β-tubulin constructs with α-CTTs, β-CTTs, or both from various human tubulin isotypes, we probed their interactions with VDAC reconstituted into planar lipid bilayers. A comparative study of the blockage kinetics revealed that either α-CTTs or β-CTTs block the VDAC pore and that the efficiency of blockage by individual CTTs spans 2 orders of magnitude, depending on the CTT isotype. β-Tubulin constructs, notably β3, blocked VDAC most effectively. We quantitatively described these experimental results using a physical model that accounted only for the number and distribution of charges in the CTT, and not for the interactions between specific residues on the CTT and VDAC pore. Based on these results, we speculate that the effectiveness of VDAC regulation by tubulin depends on the predominant tubulin isotype in a cell. Consequently, the fluxes of ATP/ADP through the channel could vary significantly, depending on the isotype, thus suggesting an intriguing link between VDAC regulation and the diversity of tubulin isotypes present in vertebrates.
Keywords: C-terminal tail; VDAC; beta-barrel channel; drift-diffusion model; engineered recombinant tubulin; gating; gating charge; intrinsically disordered protein domains; ion channel; membrane protein; membrane transport; mitochondrial transport; peripheral membrane proteins; permeability; protein engineering; protein-lipid interaction; recombinant protein expression; tubulin.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
