

The BCKDH Kinase and Phosphatase Integrate BCAA and Lipid Metabolism via Regulation of ATP-Citrate Lyase
- PMID: 29779826
- PMCID: PMC5990471
- DOI: 10.1016/j.cmet.2018.04.015
The BCKDH Kinase and Phosphatase Integrate BCAA and Lipid Metabolism via Regulation of ATP-Citrate Lyase
Abstract
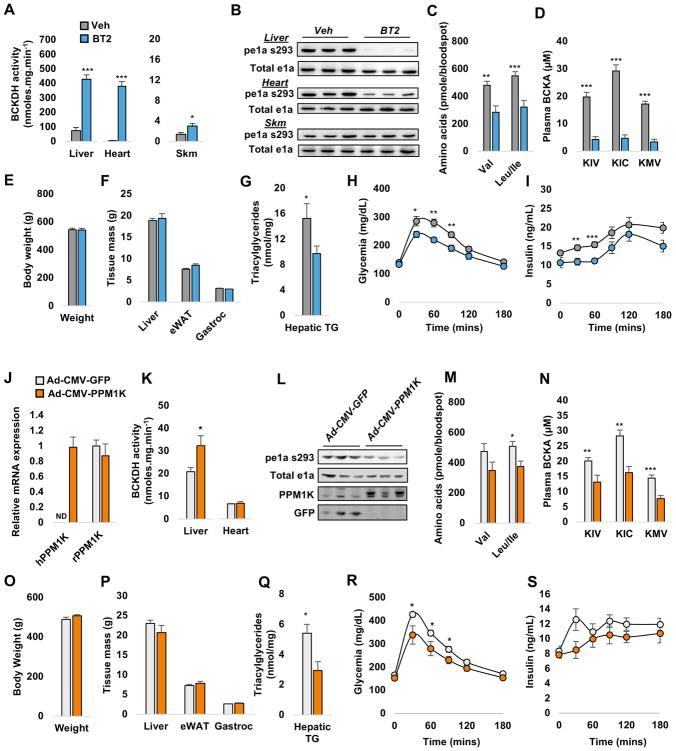
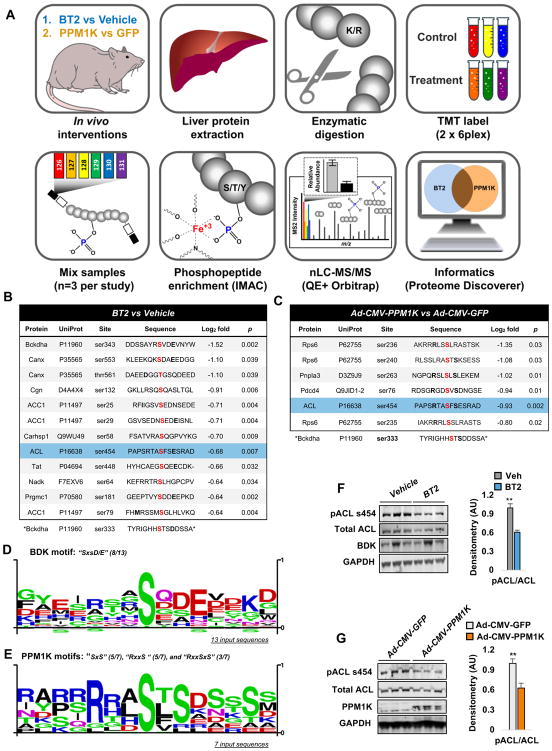
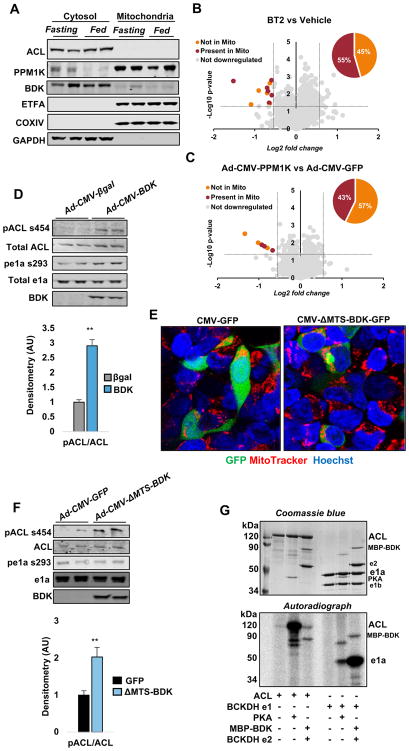
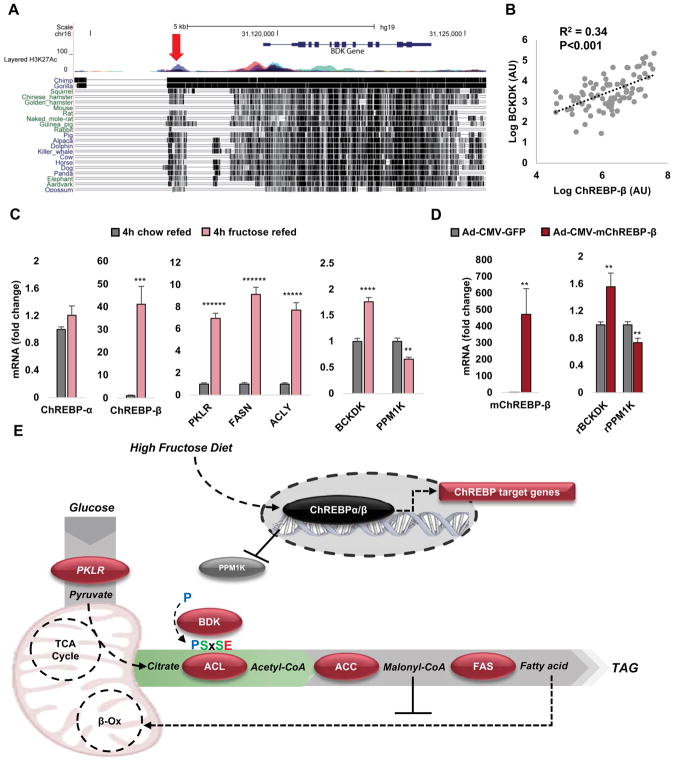
Branched-chain amino acids (BCAA) are strongly associated with dysregulated glucose and lipid metabolism, but the underlying mechanisms are poorly understood. We report that inhibition of the kinase (BDK) or overexpression of the phosphatase (PPM1K) that regulates branched-chain ketoacid dehydrogenase (BCKDH), the committed step of BCAA catabolism, lowers circulating BCAA, reduces hepatic steatosis, and improves glucose tolerance in the absence of weight loss in Zucker fatty rats. Phosphoproteomics analysis identified ATP-citrate lyase (ACL) as an alternate substrate of BDK and PPM1K. Hepatic overexpression of BDK increased ACL phosphorylation and activated de novo lipogenesis. BDK and PPM1K transcript levels were increased and repressed, respectively, in response to fructose feeding or expression of the ChREBP-β transcription factor. These studies identify BDK and PPM1K as a ChREBP-regulated node that integrates BCAA and lipid metabolism. Moreover, manipulation of the BDK:PPM1K ratio relieves key metabolic disease phenotypes in a genetic model of severe obesity.
Keywords: ATP-citrate lyase; branched-chain amino acids; diabetes; lipid metabolism; obesity; systems physiology.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
C.B.N. is a member of the Lilly Global Diabetes Advisory Board.
Figures

References
-
- Berwick DC, Hers I, Heesom KJ, Moule SK, Tavare JM. The identification of ATP-citrate lyase as a protein kinase B (Akt) substrate in primary adipocytes. J Biol Chem. 2002;277:33895–33900. - PubMed
-
- Davie JR, Wynn RM, Meng M, Huang YS, Aalund G, Chuang DT, Lau KS. Expression and characterization of branched-chain alpha-ketoacid dehydrogenase kinase from the rat. Is it a histidine-protein kinase? J Biol Chem. 1995;270:19861–19867. - PubMed
-
- DeBalsi KL, Wong KE, Koves TR, Slentz DH, Seiler SE, Wittmann AH, Ilkayeva OR, Stevens RD, Perry CGR, Lark DS, et al. Targeted metabolomics connects thioredoxin-interacting protein (TXNIP) to mitochondrial fuel selection and regulation of specific oxidoreductase enzymes in skeletal muscle. J Biol Chem. 2014;289:8106–8120. - PMC - PubMed
-
- Felig P, Marliss E, Cahill GF. Plasma amino acid levels and insulin secretion in obesity. N Engl J Med. 1969;281:811–816. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
