

The Eukaryotic Proteome Is Shaped by E3 Ubiquitin Ligases Targeting C-Terminal Degrons
- PMID: 29779948
- PMCID: PMC6003881
- DOI: 10.1016/j.cell.2018.04.028
The Eukaryotic Proteome Is Shaped by E3 Ubiquitin Ligases Targeting C-Terminal Degrons
Abstract
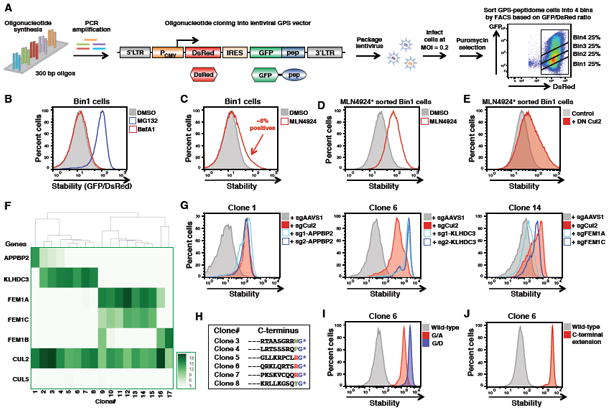
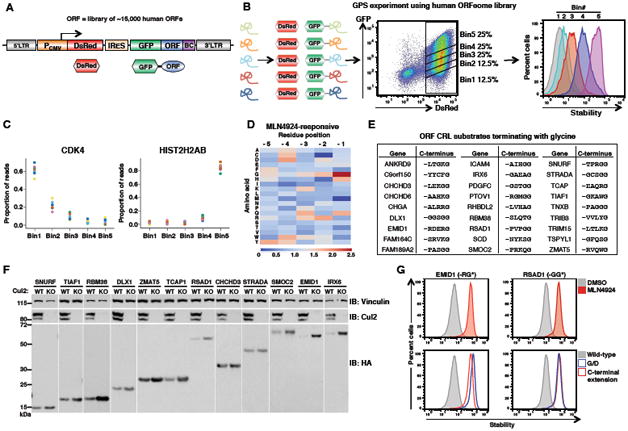
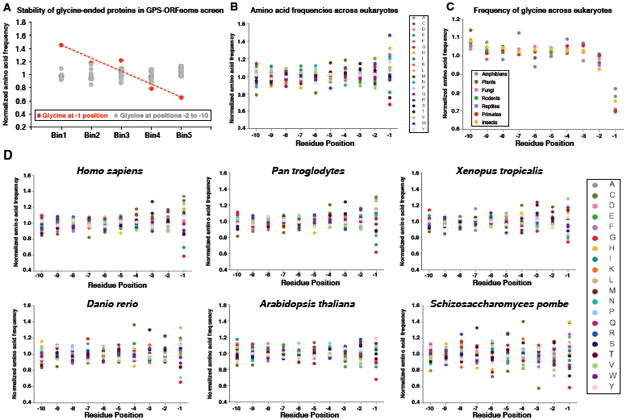
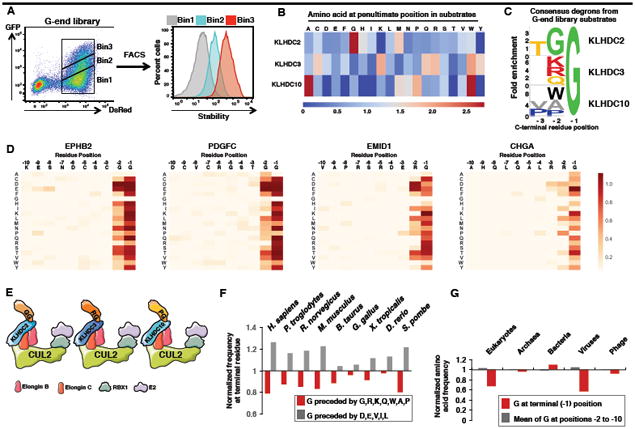
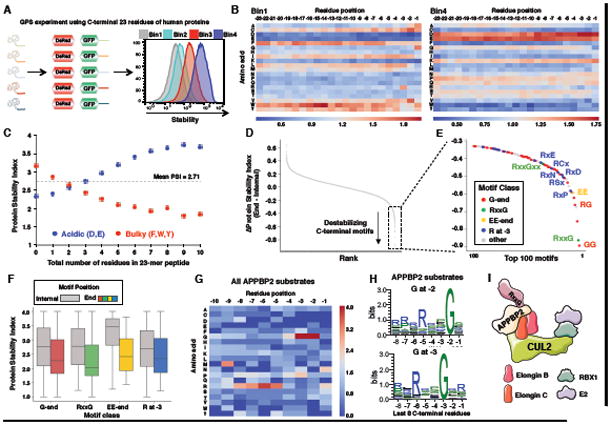
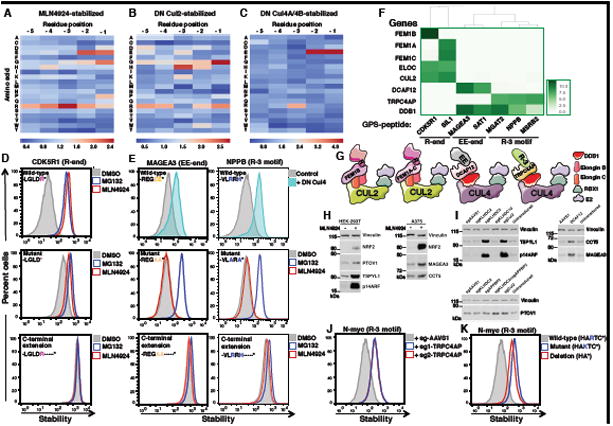
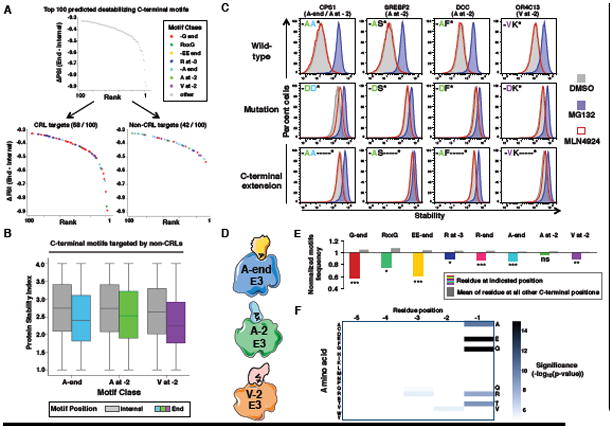
Degrons are minimal elements that mediate the interaction of proteins with degradation machineries to promote proteolysis. Despite their central role in proteostasis, the number of known degrons remains small, and a facile technology to characterize them is lacking. Using a strategy combining global protein stability (GPS) profiling with a synthetic human peptidome, we identify thousands of peptides containing degron activity. Employing CRISPR screening, we establish that the stability of many proteins is regulated through degrons located at their C terminus. We characterize eight Cullin-RING E3 ubiquitin ligase (CRL) complex adaptors that regulate C-terminal degrons, including six CRL2 and two CRL4 complexes, and computationally implicate multiple non-CRLs in end recognition. Proteome analysis revealed that the C termini of eukaryotic proteins are depleted for C-terminal degrons, suggesting an E3-ligase-dependent modulation of proteome composition. Thus, we propose that a series of "C-end rules" operate to govern protein stability and shape the eukaryotic proteome.
Keywords: C terminus; CRL; Cullin; DesCEND; E3 ubiquitin ligase; GPS; degron; global protein stability; protein degradation; ubiquitination.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
Comment in
-
At Long Last, a C-Terminal Bookend for the Ubiquitin Code.Mol Cell. 2018 May 17;70(4):568-571. doi: 10.1016/j.molcel.2018.05.006. Mol Cell. 2018. PMID: 29775575
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
