

Group I Paks support muscle regeneration and counteract cancer-associated muscle atrophy
- PMID: 29781585
- PMCID: PMC6104114
- DOI: 10.1002/jcsm.12303
Group I Paks support muscle regeneration and counteract cancer-associated muscle atrophy
Abstract
Background: Skeletal muscle is characterized by an efficient regeneration potential that is often impaired during myopathies. Understanding the molecular players involved in muscle homeostasis and regeneration could help to find new therapies against muscle degenerative disorders. Previous studies revealed that the Ser/Thr kinase p21 protein-activated kinase 1 (Pak1) was specifically down-regulated in the atrophying gastrocnemius of Yoshida hepatoma-bearing rats. In this study, we evaluated the role of group I Paks during cancer-related atrophy and muscle regeneration.
Methods: We examined Pak1 expression levels in the mouse Tibialis Anterior muscles during cancer cachexia induced by grafting colon adenocarcinoma C26 cells and in vitro by dexamethasone treatment. We investigated whether the overexpression of Pak1 counteracts muscle wasting in C26-bearing mice and in vitro also during interleukin-6 (IL6)-induced or dexamethasone-induced C2C12 atrophy. Moreover, we analysed the involvement of group I Paks on myogenic differentiation in vivo and in vitro using the group I chemical inhibitor IPA-3.
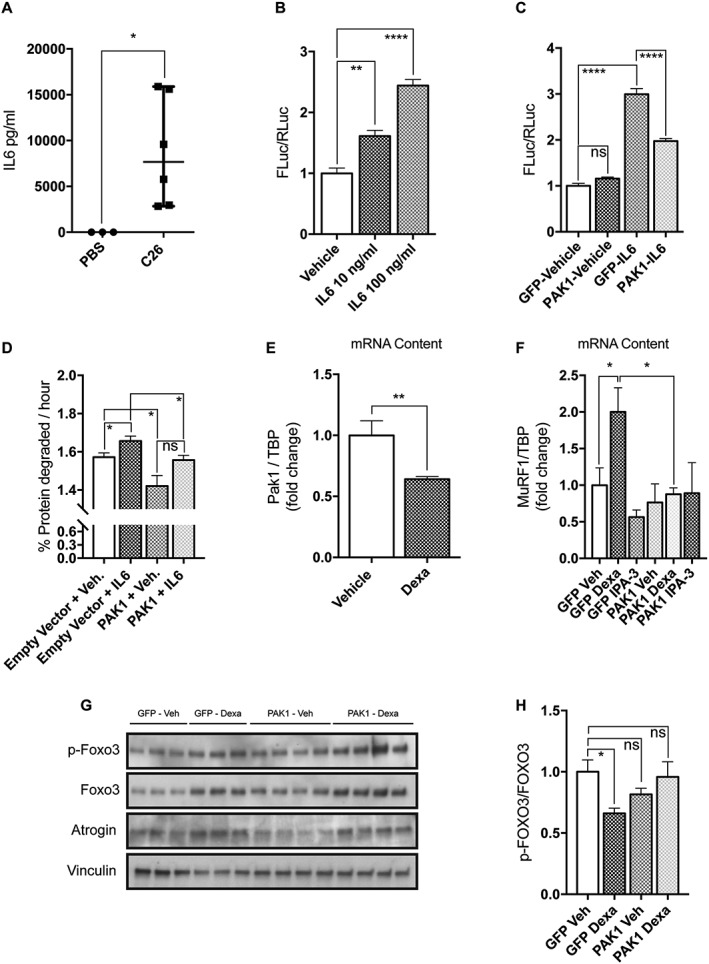
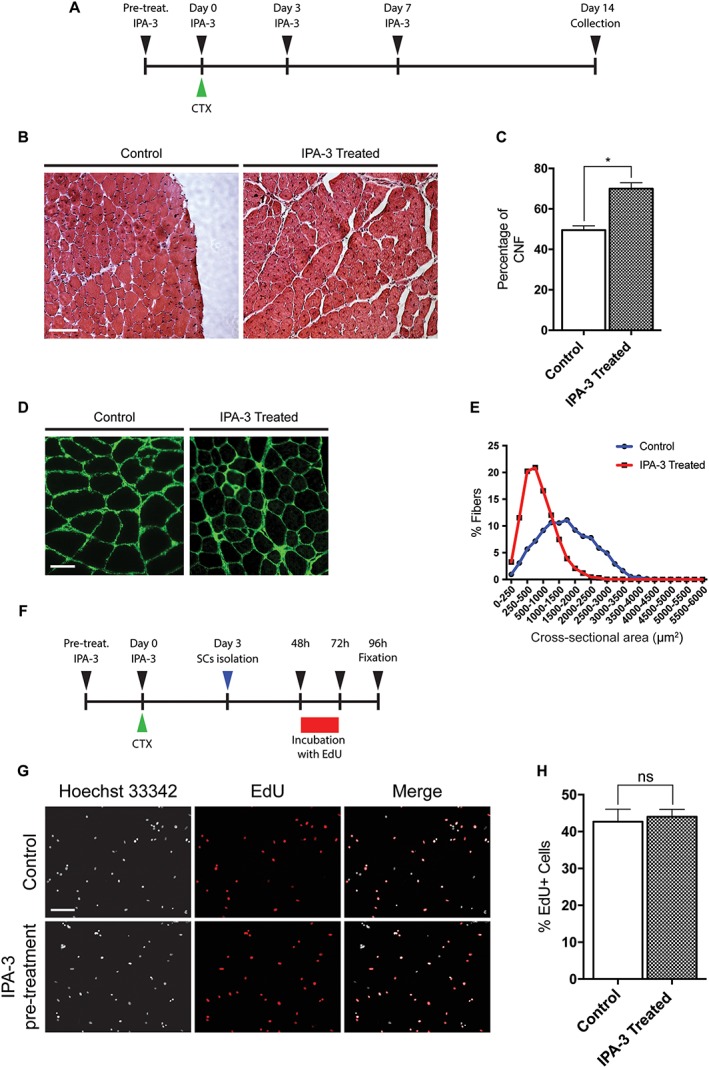
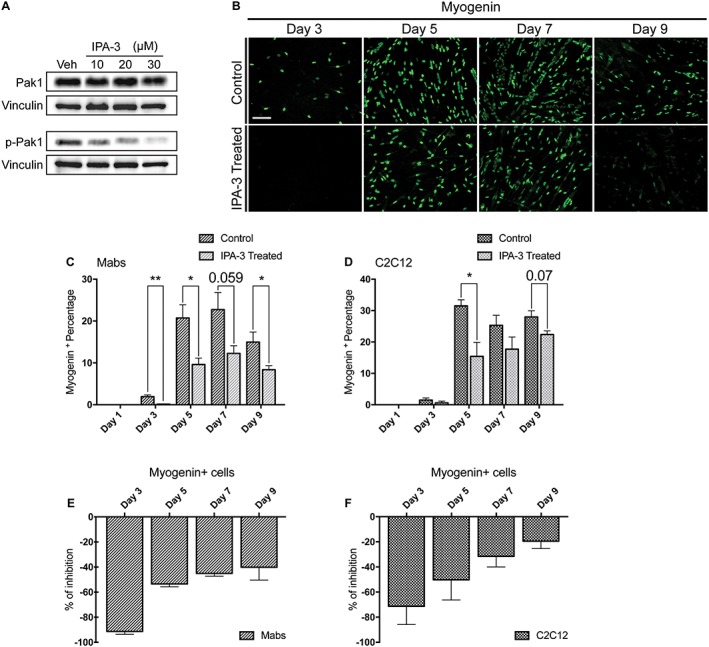
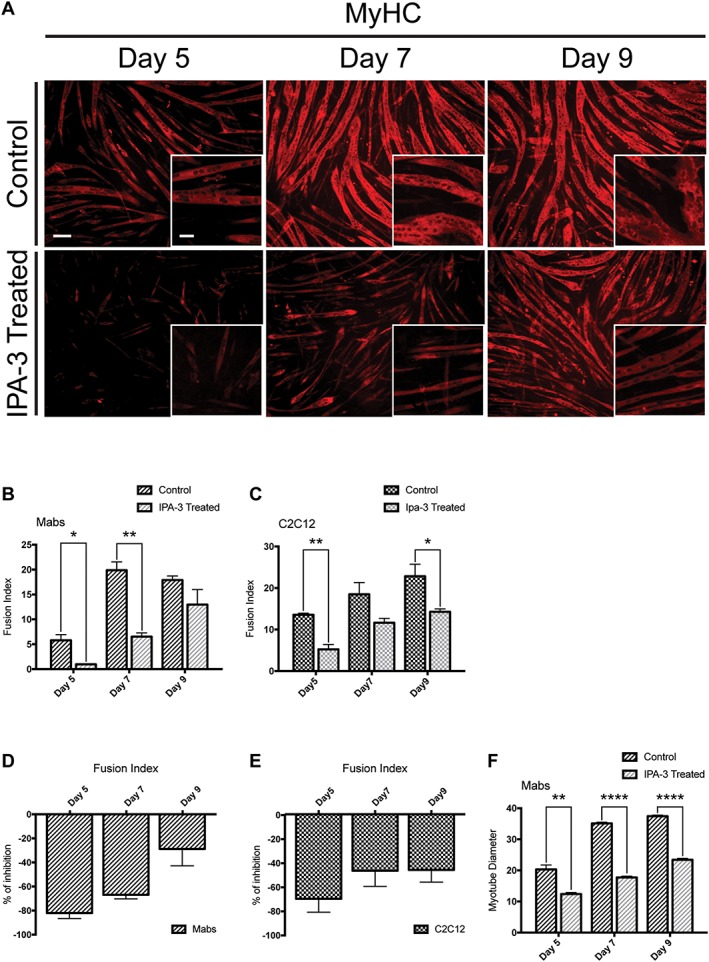
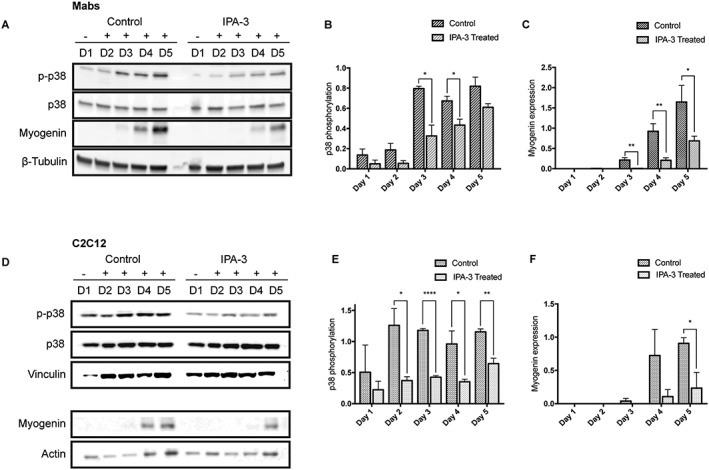
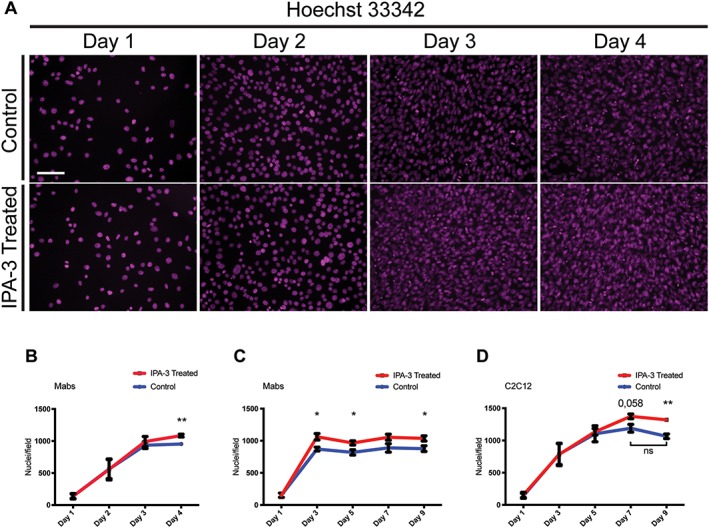
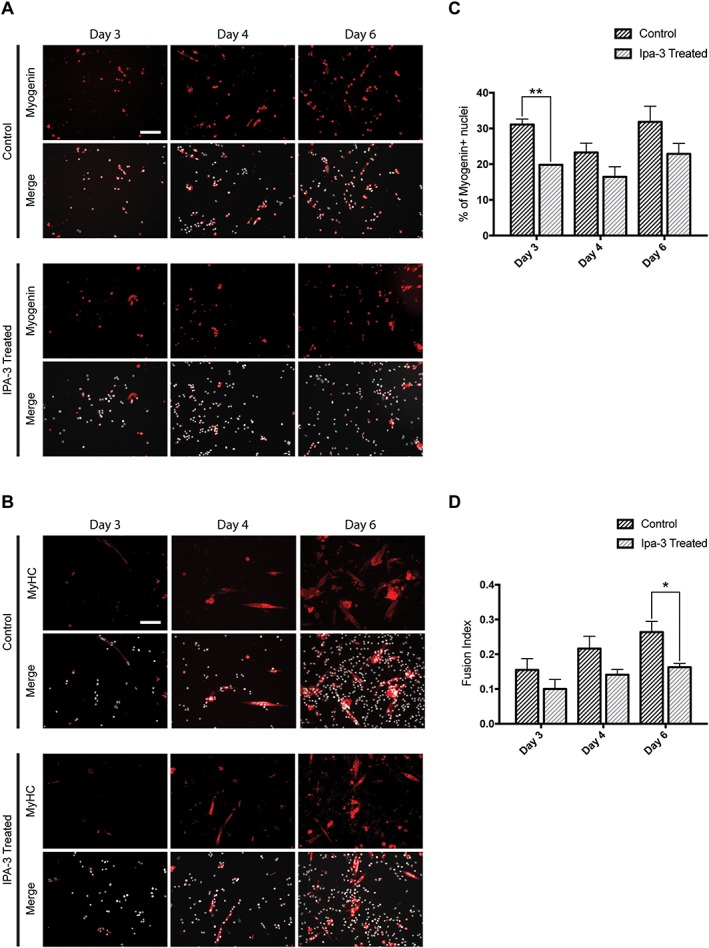
Results: We found that Pak1 expression levels are reduced during cancer-induced cachexia in the Tibialis Anterior muscles of colon adenocarcinoma C26-bearing mice and in vitro during dexamethasone-induced myotube atrophy. Electroporation of muscles of C26-bearing mice with plasmids directing the synthesis of PAK1 preserves fiber size in cachectic muscles by restraining the expression of atrogin-1 and MuRF1 and possibly by inducing myogenin expression. Consistently, the overexpression of PAK1 reduces the dexamethasone-induced expression of MuRF1 in myotubes and increases the phospho-FOXO3/FOXO3 ratio. Interestingly, the ectopic expression of PAK1 counteracts atrophy in vitro by restraining the IL6-Stat3 signalling pathway measured in luciferase-based assays and by reducing rates of protein degradation in atrophying myotubes exposed to IL6. On the other hand, we observed that the inhibition of group I Paks has no effect on myotube atrophy in vitro and is associated with impaired muscle regeneration in vivo and in vitro. In fact, we found that mice treated with the group I inhibitor IPA-3 display a delayed recovery from cardiotoxin-induced muscle injury. This is consistent with in vitro experiments showing that IPA-3 impairs myogenin expression and myotube formation in vessel-associated myogenic progenitors, C2C12 myoblasts, and satellite cells. Finally, we observed that IPA-3 reduces p38α/β phosphorylation that is required to proceed through various stages of satellite cells differentiation: activation, asymmetric division, and ultimately myotube formation.
Conclusions: Our data provide novel evidence that is consistent with group I Paks playing a central role in the regulation of muscle homeostasis, atrophy and myogenesis.
Keywords: Atrophy; Cachexia; Muscle; Paks; Regeneration.
© 2018 The Authors. Journal of Cachexia, Sarcopenia and Muscle published by John Wiley & Sons Ltd on behalf of the Society on Sarcopenia, Cachexia and Wasting Disorders.
Figures

References
-
- Wigmore SJ, Plester CE, Ross JA, Fearon KC. Contribution of anorexia and hypermetabolism to weight loss in anicteric patients with pancreatic cancer. Br J Surg 1997;84:196–197. - PubMed
-
- Fearon K, Strasser F, Anker SD, Bosaeus I, Bruera E, Fainsinger RL, et al. Definition and classification of cancer cachexia: an international consensus. Lancet Oncol Elsevier 2011;12:489–495. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
