

Endoplasmic reticulum-plasma membrane contact sites integrate sterol and phospholipid regulation
- PMID: 29782498
- PMCID: PMC5983861
- DOI: 10.1371/journal.pbio.2003864
Endoplasmic reticulum-plasma membrane contact sites integrate sterol and phospholipid regulation
Abstract
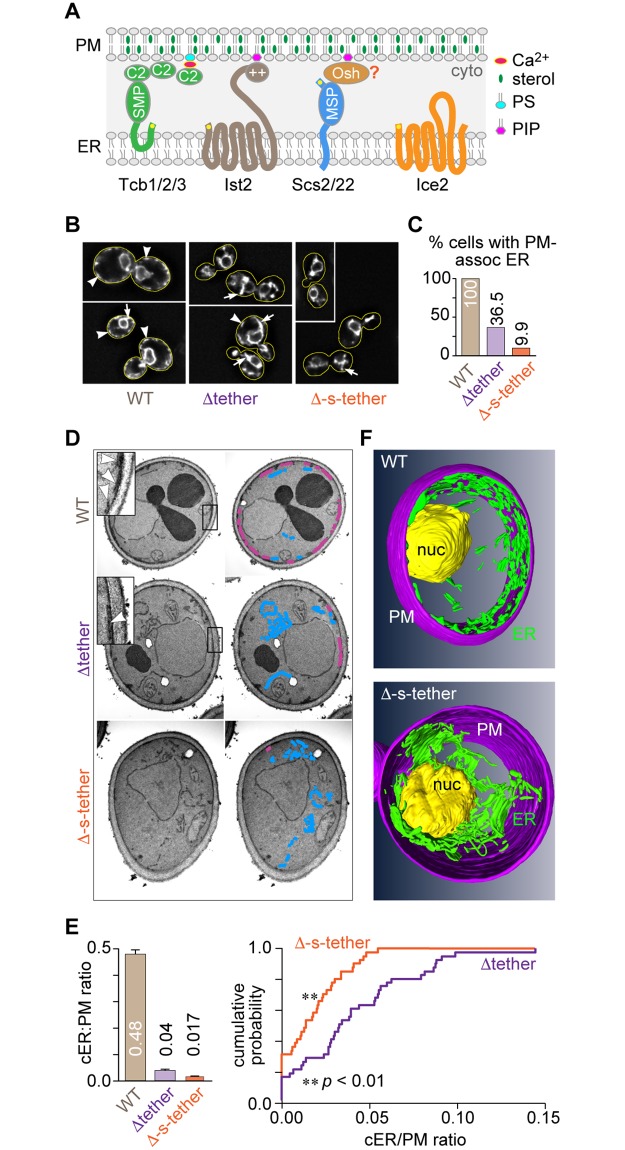
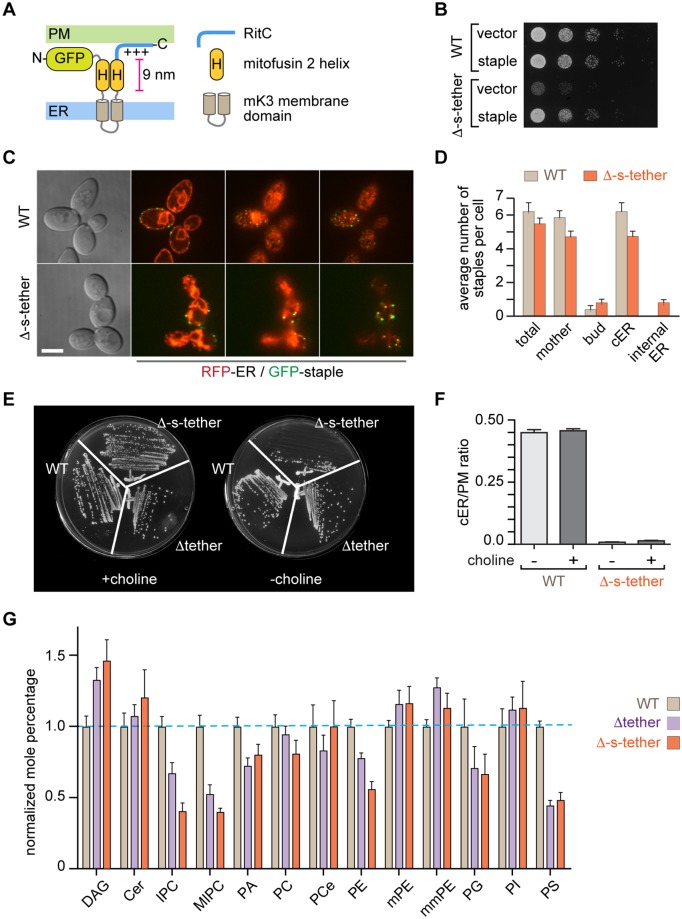
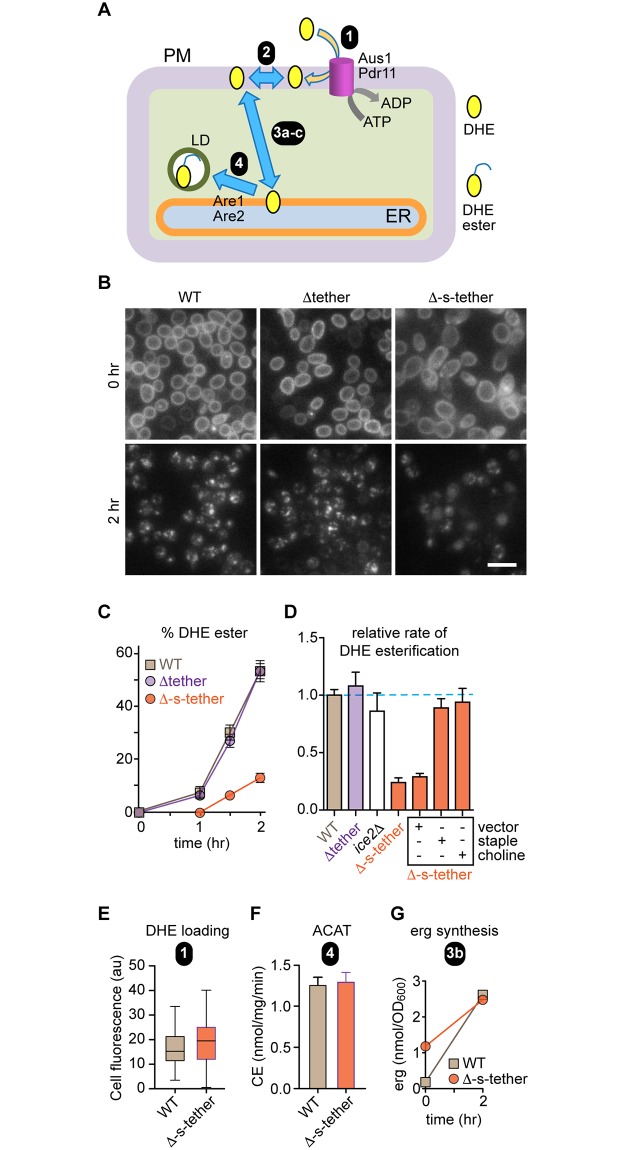
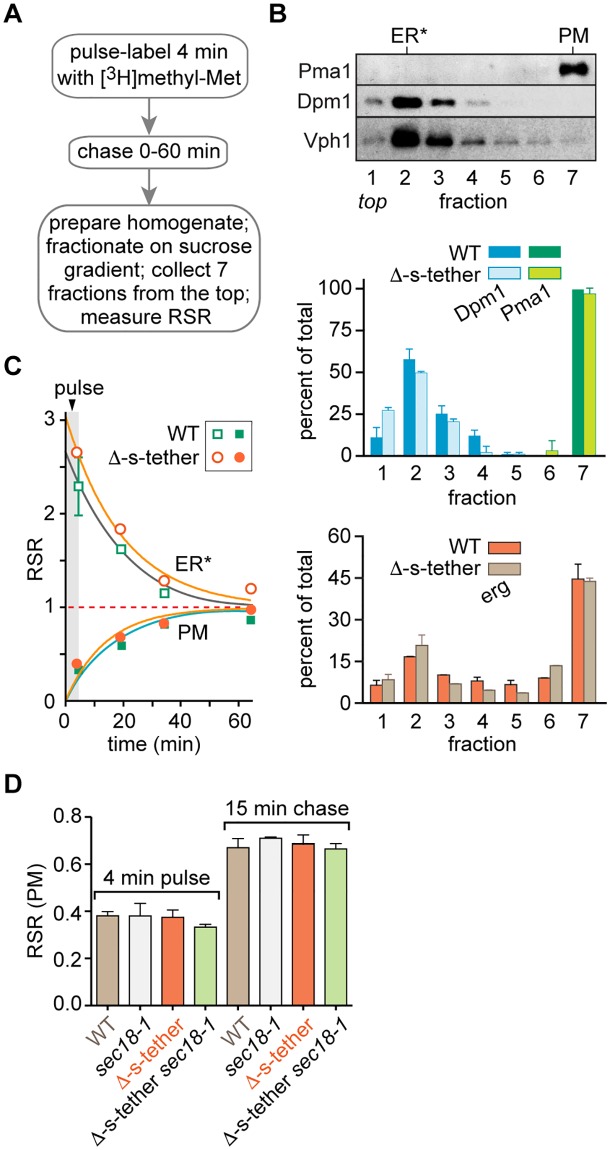
Tether proteins attach the endoplasmic reticulum (ER) to other cellular membranes, thereby creating contact sites that are proposed to form platforms for regulating lipid homeostasis and facilitating non-vesicular lipid exchange. Sterols are synthesized in the ER and transported by non-vesicular mechanisms to the plasma membrane (PM), where they represent almost half of all PM lipids and contribute critically to the barrier function of the PM. To determine whether contact sites are important for both sterol exchange between the ER and PM and intermembrane regulation of lipid metabolism, we generated Δ-super-tether (Δ-s-tether) yeast cells that lack six previously identified tethering proteins (yeast extended synatotagmin [E-Syt], vesicle-associated membrane protein [VAMP]-associated protein [VAP], and TMEM16-anoctamin homologues) as well as the presumptive tether Ice2. Despite the lack of ER-PM contacts in these cells, ER-PM sterol exchange is robust, indicating that the sterol transport machinery is either absent from or not uniquely located at contact sites. Unexpectedly, we found that the transport of exogenously supplied sterol to the ER occurs more slowly in Δ-s-tether cells than in wild-type (WT) cells. We pinpointed this defect to changes in sterol organization and transbilayer movement within the PM bilayer caused by phospholipid dysregulation, evinced by changes in the abundance and organization of PM lipids. Indeed, deletion of either OSH4, which encodes a sterol/phosphatidylinositol-4-phosphate (PI4P) exchange protein, or SAC1, which encodes a PI4P phosphatase, caused synthetic lethality in Δ-s-tether cells due to disruptions in redundant PI4P and phospholipid regulatory pathways. The growth defect of Δ-s-tether cells was rescued with an artificial "ER-PM staple," a tether assembled from unrelated non-yeast protein domains, indicating that endogenous tether proteins have nonspecific bridging functions. Finally, we discovered that sterols play a role in regulating ER-PM contact site formation. In sterol-depleted cells, levels of the yeast E-Syt tether Tcb3 were induced and ER-PM contact increased dramatically. These results support a model in which ER-PM contact sites provide a nexus for coordinating the complex interrelationship between sterols, sphingolipids, and phospholipids that maintain PM composition and integrity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Holthuis JC, Levine TP. Lipid traffic: floppy drives and a superhighway. Nat Rev Mol Cell Biol. 2005;6(3):209–20. Epub 2005/03/02. doi: 10.1038/nrm1591 . - DOI - PubMed
-
- Lev S. Non-vesicular lipid transport by lipid-transfer proteins and beyond. Nat Rev Mol Cell Biol. 2010;11(10):739–50. Epub 2010/09/09. doi: 10.1038/nrm2971 . - DOI - PubMed
-
- Holthuis JC, Menon AK. Lipid landscapes and pipelines in membrane homeostasis. Nature. 2014;510(7503):48–57. Epub 2014/06/06. doi: 10.1038/nature13474 . - DOI - PubMed
-
- Pichler H, Gaigg B, Hrastnik C, Achleitner G, Kohlwein SD, Zellnig G, et al. A subfraction of the yeast endoplasmic reticulum associates with the plasma membrane and has a high capacity to synthesize lipids. Eur J Biochem. 2001;268(8):2351–61. . - PubMed
-
- Manford AG, Stefan CJ, Yuan HL, Macgurn JA, Emr SD. ER-to-plasma membrane tethering proteins regulate cell signaling and ER morphology. Dev Cell. 2012;23(6):1129–40. Epub 2012/12/15. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
