

Short-chain dehydrogenase/reductase governs steroidal specialized metabolites structural diversity and toxicity in the genus Solanum
- PMID: 29784829
- PMCID: PMC6003347
- DOI: 10.1073/pnas.1804835115
Short-chain dehydrogenase/reductase governs steroidal specialized metabolites structural diversity and toxicity in the genus Solanum
Abstract
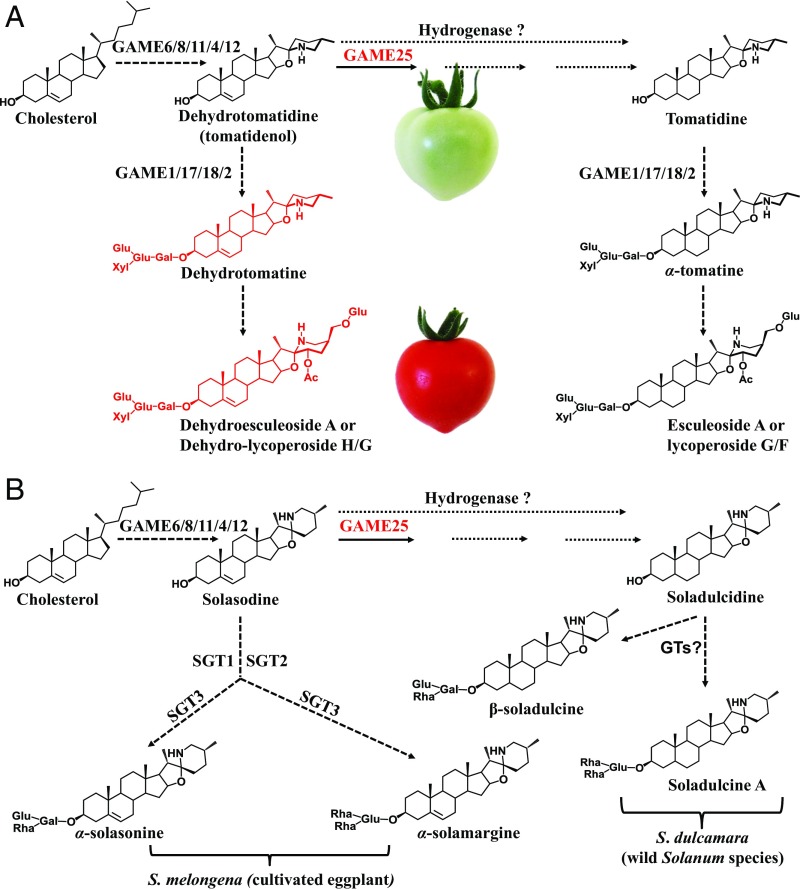
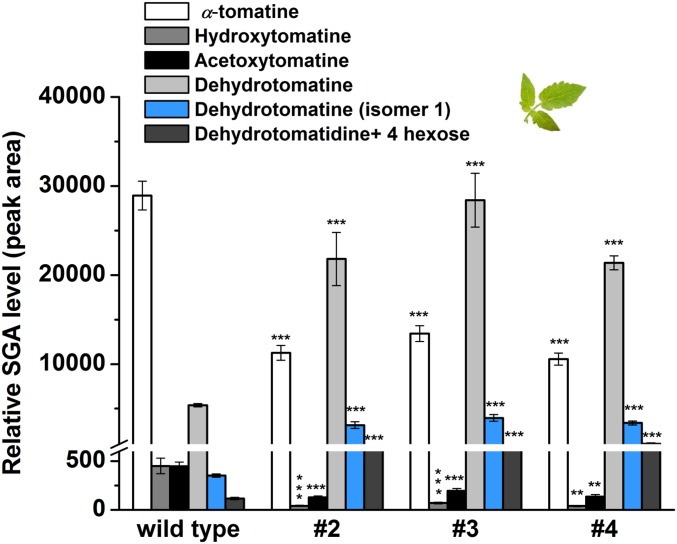
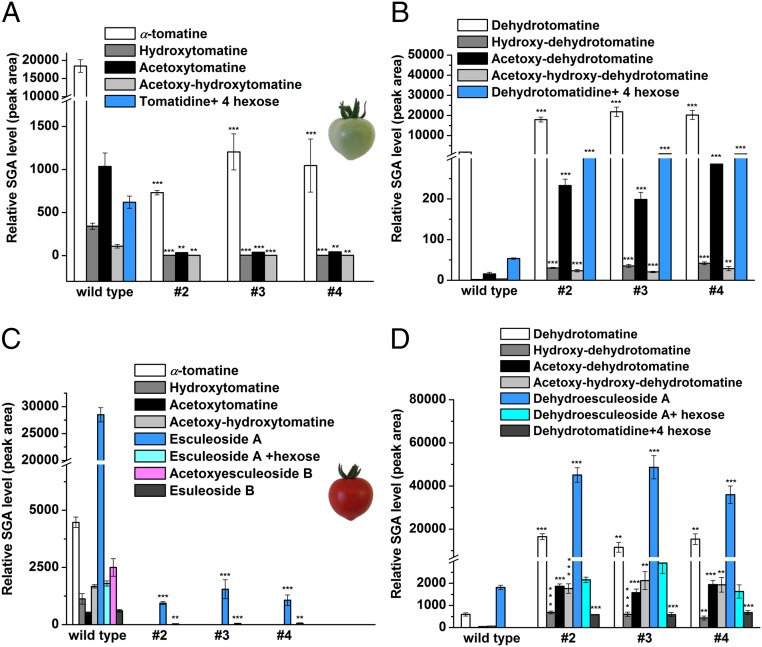
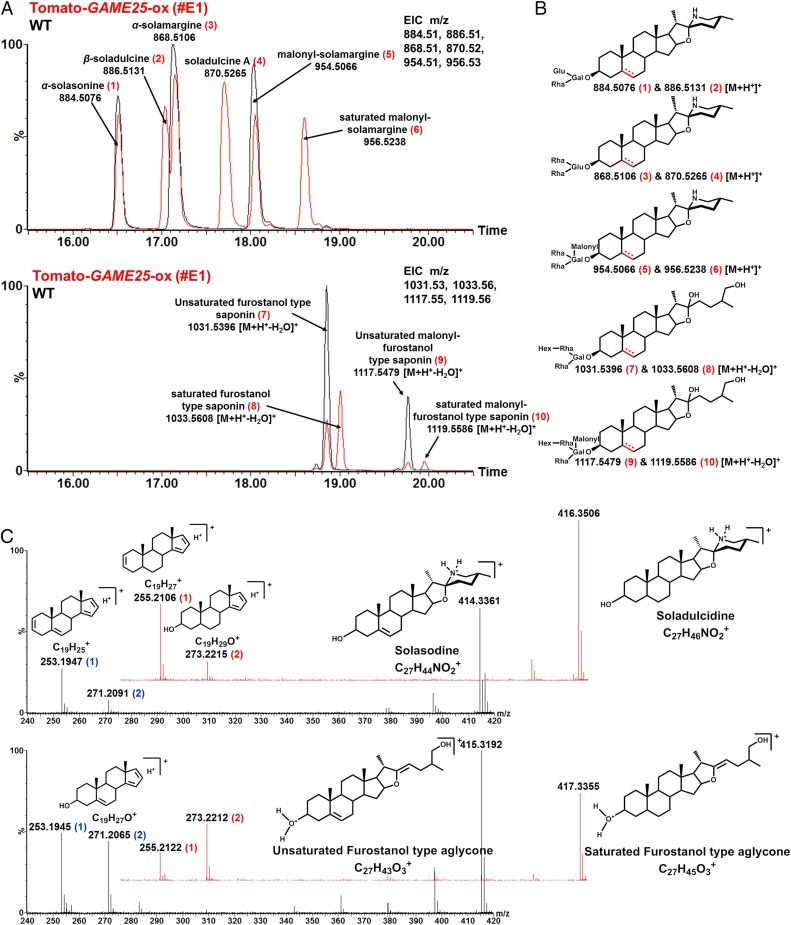
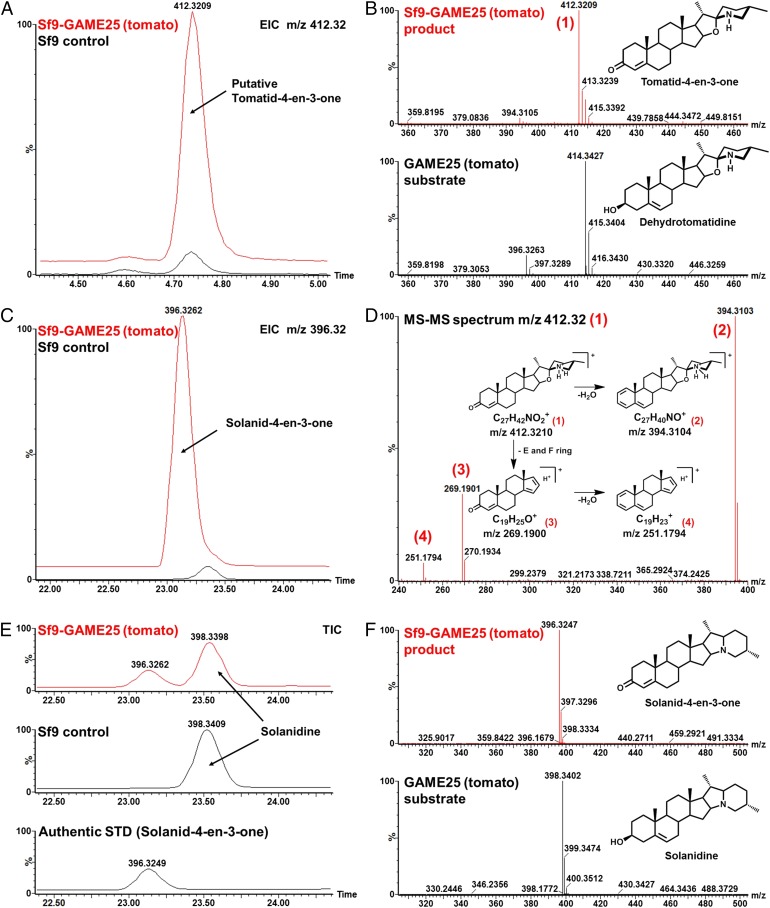
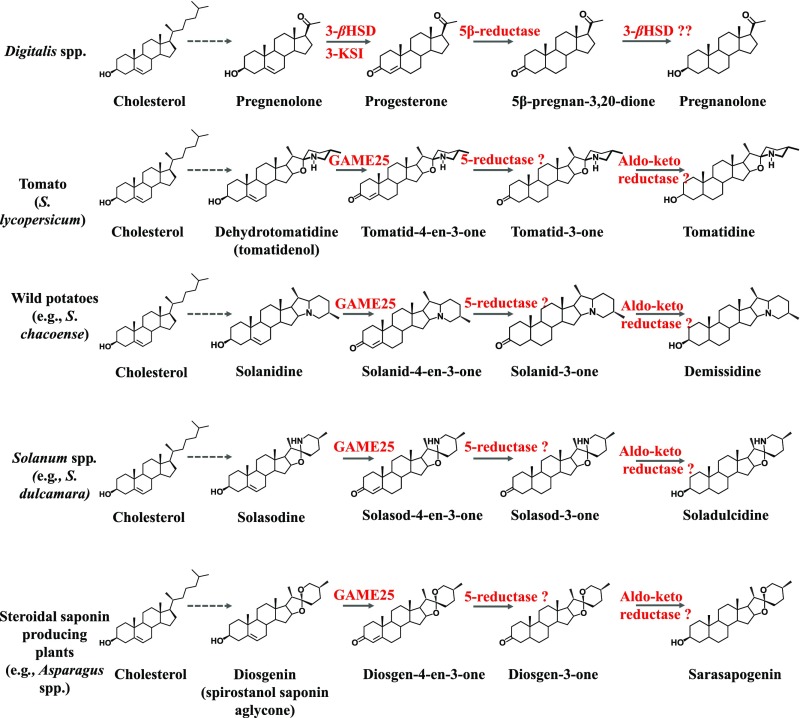
Thousands of specialized, steroidal metabolites are found in a wide spectrum of plants. These include the steroidal glycoalkaloids (SGAs), produced primarily by most species of the genus Solanum, and metabolites belonging to the steroidal saponins class that are widespread throughout the plant kingdom. SGAs play a protective role in plants and have potent activity in mammals, including antinutritional effects in humans. The presence or absence of the double bond at the C-5,6 position (unsaturated and saturated, respectively) creates vast structural diversity within this metabolite class and determines the degree of SGA toxicity. For many years, the elimination of the double bond from unsaturated SGAs was presumed to occur through a single hydrogenation step. In contrast to this prior assumption, here, we show that the tomato GLYCOALKALOID METABOLISM25 (GAME25), a short-chain dehydrogenase/reductase, catalyzes the first of three prospective reactions required to reduce the C-5,6 double bond in dehydrotomatidine to form tomatidine. The recombinant GAME25 enzyme displayed 3β-hydroxysteroid dehydrogenase/Δ5,4 isomerase activity not only on diverse steroidal alkaloid aglycone substrates but also on steroidal saponin aglycones. Notably, GAME25 down-regulation rerouted the entire tomato SGA repertoire toward the dehydro-SGAs branch rather than forming the typically abundant saturated α-tomatine derivatives. Overexpressing the tomato GAME25 in the tomato plant resulted in significant accumulation of α-tomatine in ripe fruit, while heterologous expression in cultivated eggplant generated saturated SGAs and atypical saturated steroidal saponin glycosides. This study demonstrates how a single scaffold modification of steroidal metabolites in plants results in extensive structural diversity and modulation of product toxicity.
Keywords: antinutritional; specialized metabolism; steroidal glycoalkaloids; structural diversity; tomato.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Identification of a 3β-Hydroxysteroid Dehydrogenase/ 3-Ketosteroid Reductase Involved in α-Tomatine Biosynthesis in Tomato.Plant Cell Physiol. 2019 Jun 1;60(6):1304-1315. doi: 10.1093/pcp/pcz049. Plant Cell Physiol. 2019. PMID: 30892648
-
Transcription factors SlNOR and SlNOR-like1 regulate steroidal glycoalkaloids biosynthesis in tomato fruit.Int J Biol Macromol. 2025 Apr;299:140157. doi: 10.1016/j.ijbiomac.2025.140157. Epub 2025 Jan 21. Int J Biol Macromol. 2025. PMID: 39848374
-
GLYCOALKALOID METABOLISM1 is required for steroidal alkaloid glycosylation and prevention of phytotoxicity in tomato.Plant Cell. 2011 Dec;23(12):4507-25. doi: 10.1105/tpc.111.088732. Epub 2011 Dec 16. Plant Cell. 2011. PMID: 22180624 Free PMC article.
-
Review on toxicology and activity of tomato glycoalkaloids in immature tomatoes.Food Chem. 2024 Jul 30;447:138937. doi: 10.1016/j.foodchem.2024.138937. Epub 2024 Mar 11. Food Chem. 2024. PMID: 38492295 Review.
-
The Therapeutic Value of Solanum Steroidal (Glyco)Alkaloids: A 10-Year Comprehensive Review.Molecules. 2023 Jun 23;28(13):4957. doi: 10.3390/molecules28134957. Molecules. 2023. PMID: 37446619 Free PMC article. Review.
Cited by
-
Genome-wide identification and functional characterization of borneol dehydrogenases in Wurfbainia villosa.Planta. 2023 Aug 22;258(4):69. doi: 10.1007/s00425-023-04221-0. Planta. 2023. PMID: 37608037
-
Microbial Interactions Influence the Chemical Defense of Wild and Cultivated Tomato Species.J Chem Ecol. 2025 Apr 7;51(2):47. doi: 10.1007/s10886-025-01598-y. J Chem Ecol. 2025. PMID: 40192909 Free PMC article.
-
Specialized metabolism by trichome-enriched Rubisco and fatty acid synthase components.Plant Physiol. 2023 Feb 12;191(2):1199-1213. doi: 10.1093/plphys/kiac487. Plant Physiol. 2023. PMID: 36264116 Free PMC article.
-
A High-Throughput Extraction and Analysis Method for Steroidal Glycoalkaloids in Tomato.Front Plant Sci. 2020 Jun 18;11:767. doi: 10.3389/fpls.2020.00767. eCollection 2020. Front Plant Sci. 2020. PMID: 32636855 Free PMC article.
-
Phenylpropanoid Metabolism in Astringent and Nonastringent Persimmon (Diospyros kaki) Cultivars Determines Sensitivity to Alternaria Infection.J Agric Food Chem. 2021 May 26;69(20):5628-5637. doi: 10.1021/acs.jafc.1c01312. Epub 2021 May 13. J Agric Food Chem. 2021. PMID: 33983017 Free PMC article.
References
-
- Friedman M, McDonald G, Filadelfi-keszi M. Potato glycoalkaloids: Chemistry analysis safety and plant physiology. Crit Rev Plant Sci. 1997;16:55–132.
-
- Friedman M. Tomato glycoalkaloids: Role in the plant and in the diet. J Agric Food Chem. 2002;50:5751–5780. - PubMed
-
- Friedman M. Potato glycoalkaloids and metabolites: Roles in the plant and in the diet. J Agric Food Chem. 2006;54:8655–8681. - PubMed
-
- Milner SE, et al. Bioactivities of glycoalkaloids and their aglycones from Solanum species. J Agric Food Chem. 2011;59:3454–3484. - PubMed
-
- Roddick JG. Steroidal glycoalkaloids: Nature and consequences of bioactivity. Adv Exp Med Biol. 1996;404:277–295. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
