

Resetting the epigenetic balance of Polycomb and COMPASS function at enhancers for cancer therapy
- PMID: 29785026
- PMCID: PMC6055231
- DOI: 10.1038/s41591-018-0034-6
Resetting the epigenetic balance of Polycomb and COMPASS function at enhancers for cancer therapy
Abstract
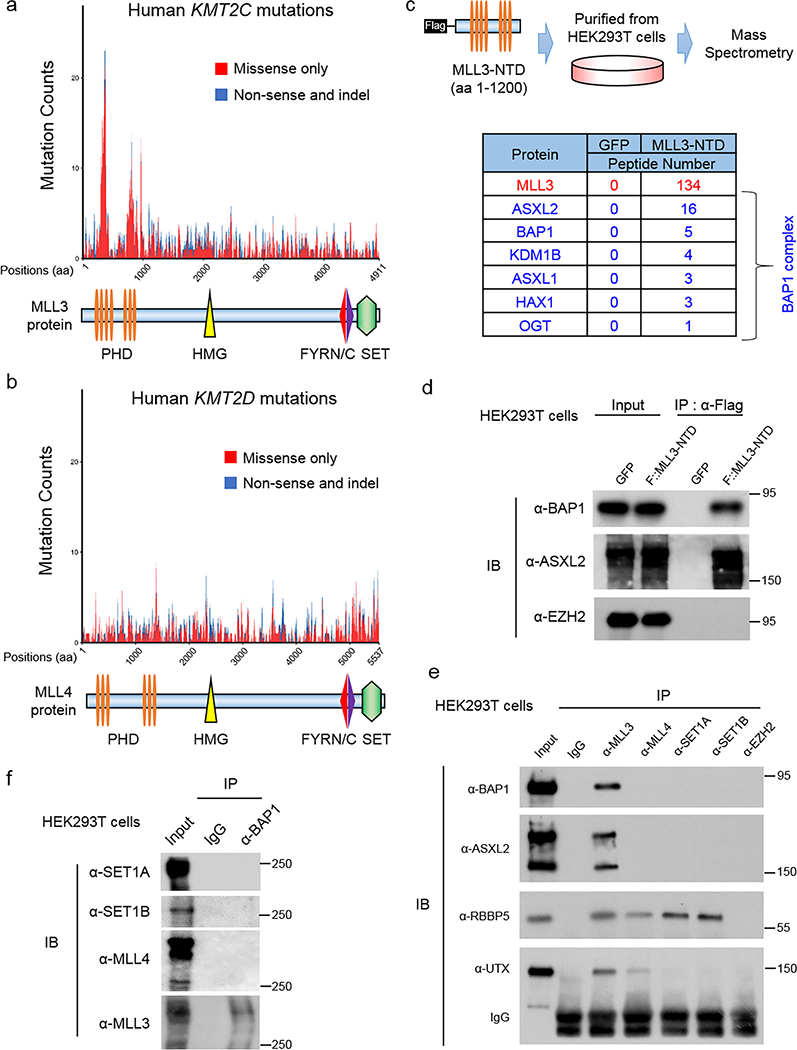
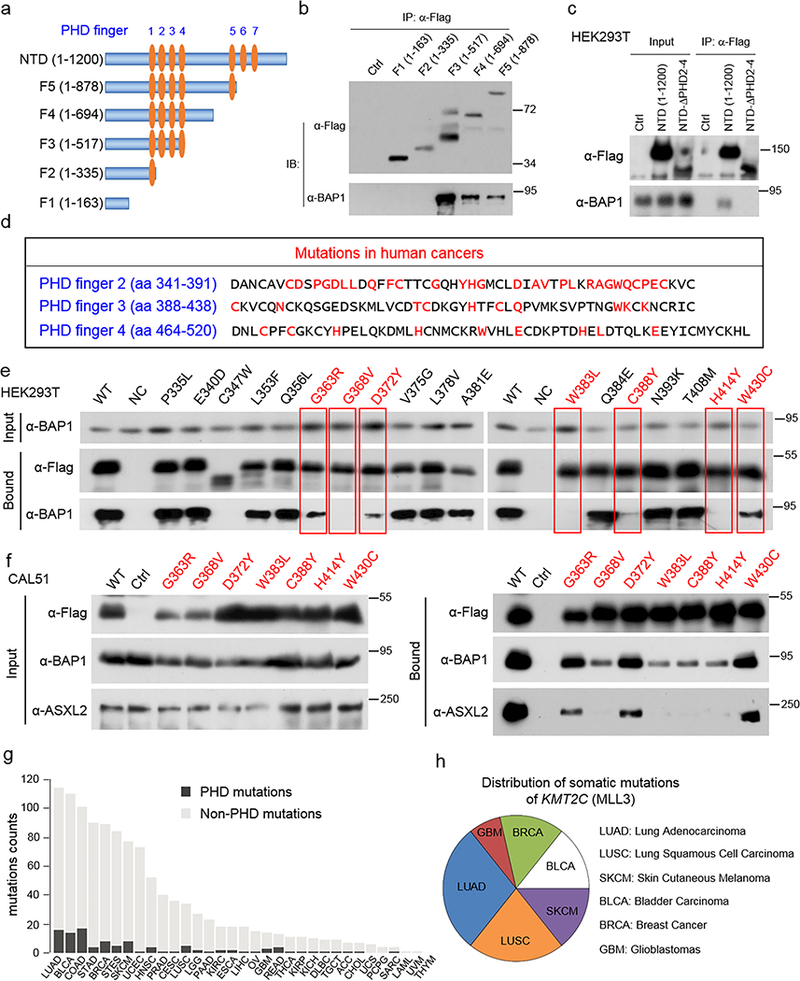
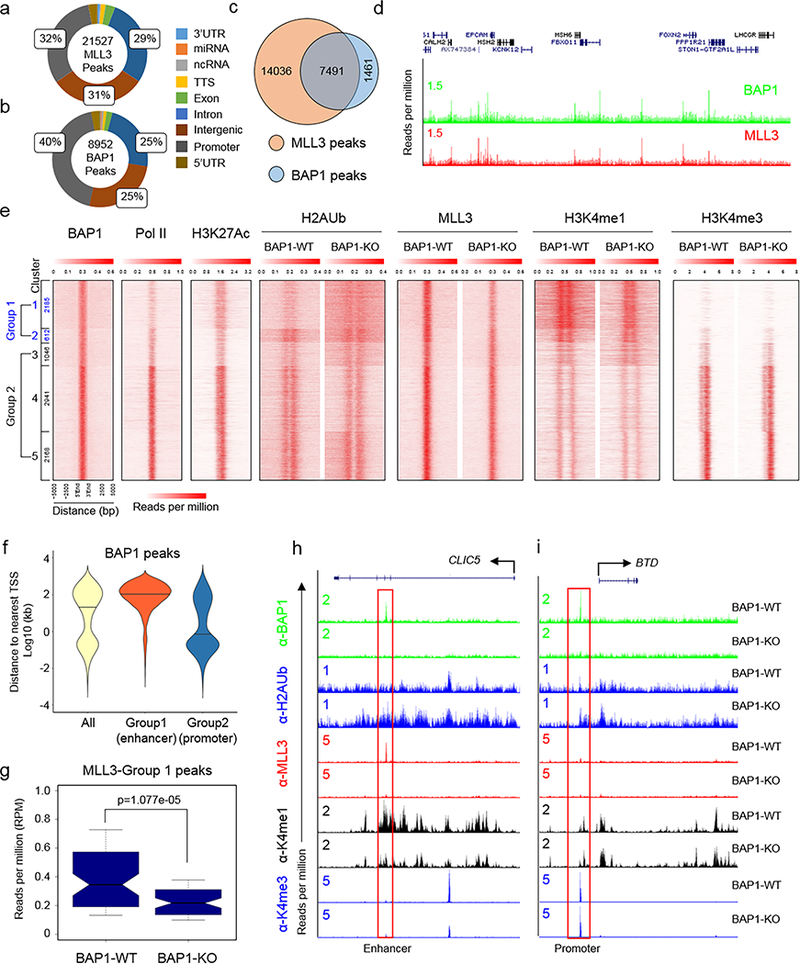
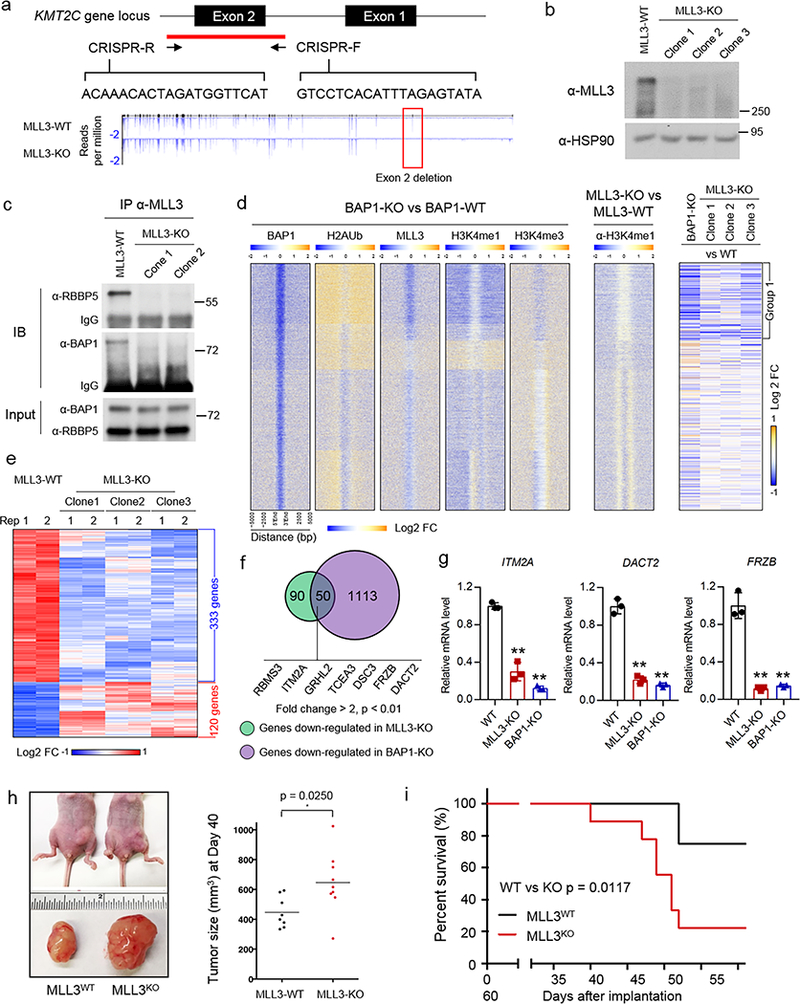
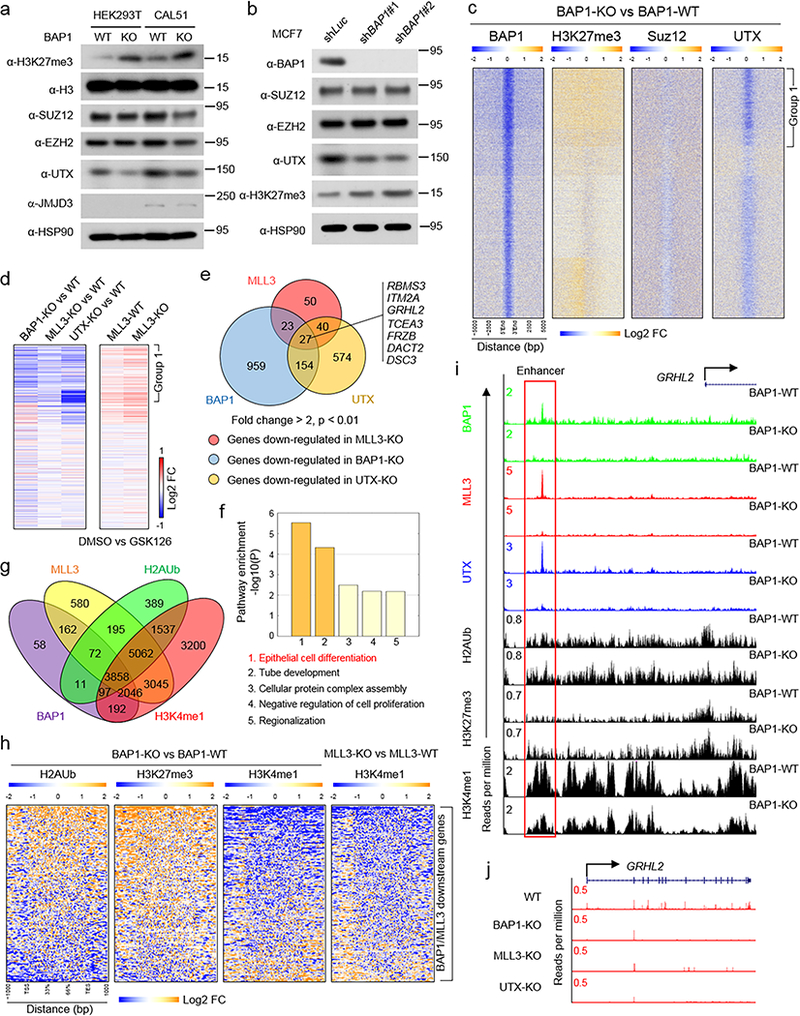
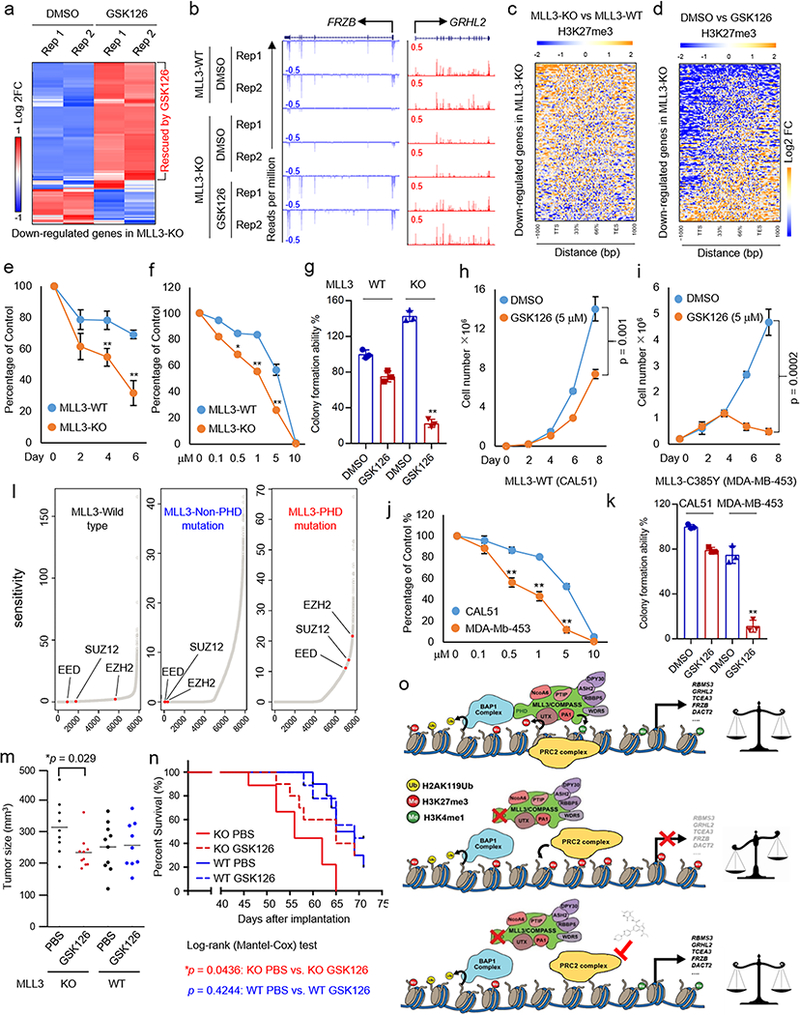
The lysine methyltransferase KMT2C (also known as MLL3), a subunit of the COMPASS complex, implements monomethylation of Lys4 on histone H3 (H3K4) at gene enhancers. KMT2C (hereafter referred to as MLL3) frequently incurs point mutations across a range of human tumor types, but precisely how these lesions alter MLL3 function and contribute to oncogenesis is unclear. Here we report a cancer mutational hotspot in MLL3 within the region encoding its plant homeodomain (PHD) repeats and demonstrate that this domain mediates association of MLL3 with the histone H2A deubiquitinase and tumor suppressor BAP1. Cancer-associated mutations in the sequence encoding the MLL3 PHD repeats disrupt the interaction between MLL3 and BAP1 and correlate with poor patient survival. Cancer cells that had PHD-associated MLL3 mutations or lacked BAP1 showed reduced recruitment of MLL3 and the H3K27 demethylase KDM6A (also known as UTX) to gene enhancers. As a result, inhibition of the H3K27 methyltransferase activity of the Polycomb repressive complex 2 (PRC2) in tumor cells harboring BAP1 or MLL3 mutations restored normal gene expression patterns and impaired cell proliferation in vivo. This study provides mechanistic insight into the oncogenic effects of PHD-associated mutations in MLL3 and suggests that restoration of a balanced state of Polycomb-COMPASS activity may have therapeutic efficacy in tumors that bear mutations in the genes encoding these epigenetic factors.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
