

Mechanisms of Action and Cell Death Associated with Clostridium perfringens Toxins
- PMID: 29786671
- PMCID: PMC5983268
- DOI: 10.3390/toxins10050212
Mechanisms of Action and Cell Death Associated with Clostridium perfringens Toxins
Abstract
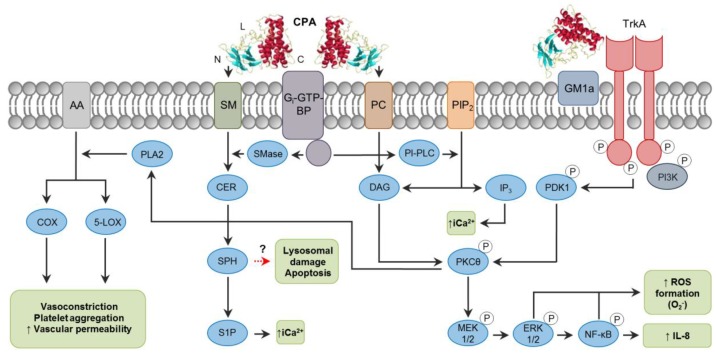
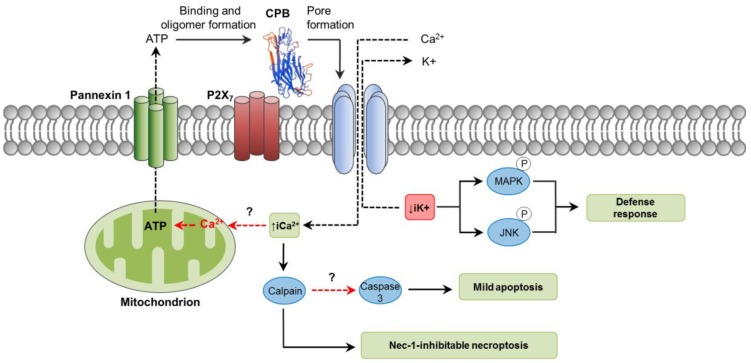
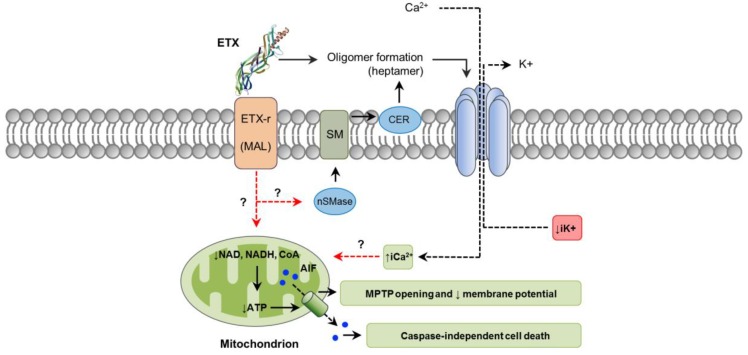
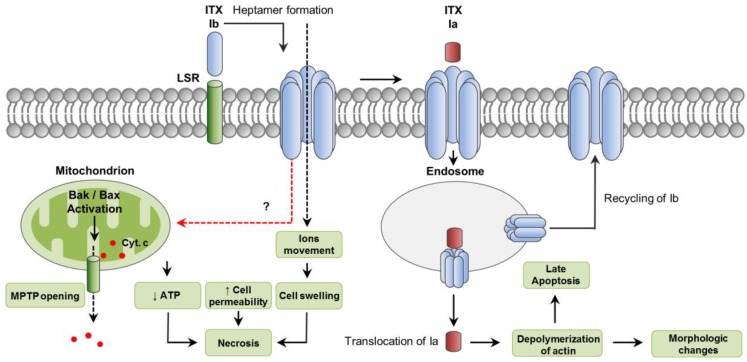
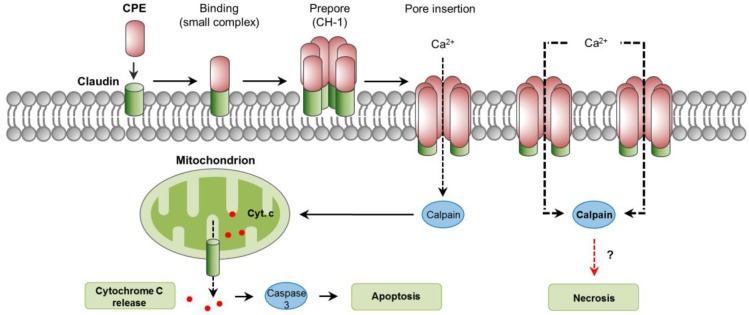
Clostridium perfringens uses its large arsenal of protein toxins to produce histotoxic, neurologic and intestinal infections in humans and animals. The major toxins involved in diseases are alpha (CPA), beta (CPB), epsilon (ETX), iota (ITX), enterotoxin (CPE), and necrotic B-like (NetB) toxins. CPA is the main virulence factor involved in gas gangrene in humans, whereas its role in animal diseases is limited and controversial. CPB is responsible for necrotizing enteritis and enterotoxemia, mostly in neonatal individuals of many animal species, including humans. ETX is the main toxin involved in enterotoxemia of sheep and goats. ITX has been implicated in cases of enteritis in rabbits and other animal species; however, its specific role in causing disease has not been proved. CPE is responsible for human food-poisoning and non-foodborne C. perfringens-mediated diarrhea. NetB is the cause of necrotic enteritis in chickens. In most cases, host⁻toxin interaction starts on the plasma membrane of target cells via specific receptors, resulting in the activation of intracellular pathways with a variety of effects, commonly including cell death. In general, the molecular mechanisms of cell death associated with C. perfringens toxins involve features of apoptosis, necrosis and/or necroptosis.
Keywords: Clostridium perfringens; apoptosis; cell death; mechanisms; necrosis; toxins.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Uzal F.A., Freedman J.C., Shrestha A., Theoret J.R., Garcia J., Awad M.M., Adams V., Moore R.J., Rood J.I., McClane B.A. Towards an understanding of the role of Clostridium perfringens toxins in human and animal disease. Future Microbiol. 2014;9:361–377. doi: 10.2217/fmb.13.168. - DOI - PMC - PubMed
-
- Revitt-Mills S.A., Rood J.I., Adams V. Clostridium Perfringens Extracellular Toxins and Enzymes: 20 and Counting. Microbiol. Aust. 2015;36:114–117. doi: 10.1071/MA15039. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
