

CLERK is a novel receptor kinase required for sensing of root-active CLE peptides in Arabidopsis
- PMID: 29789310
- PMCID: PMC6001375
- DOI: 10.1242/dev.162354
CLERK is a novel receptor kinase required for sensing of root-active CLE peptides in Arabidopsis
Abstract
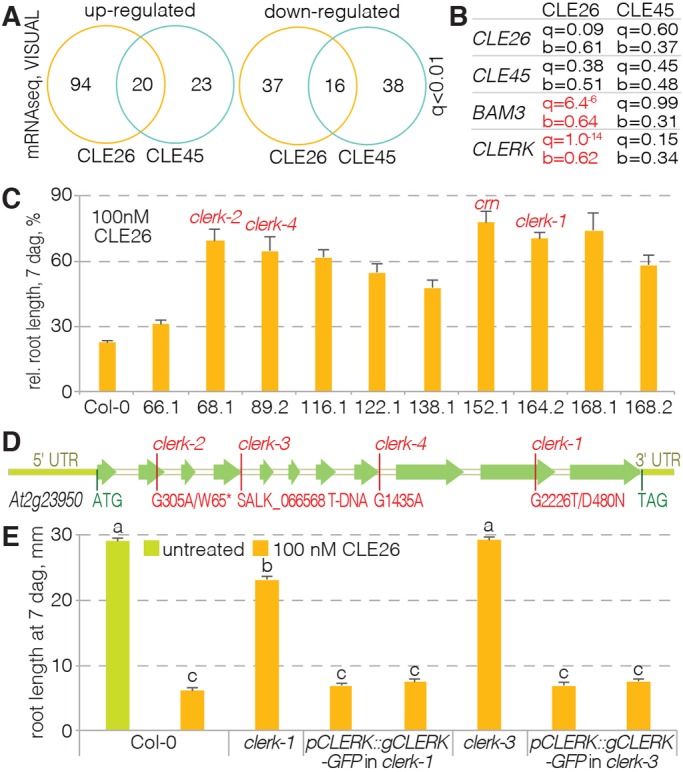
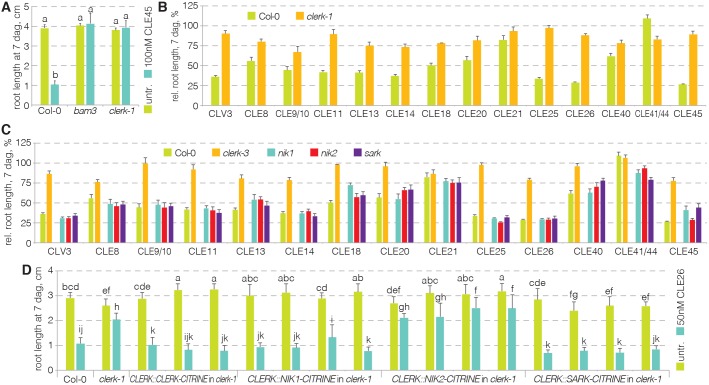
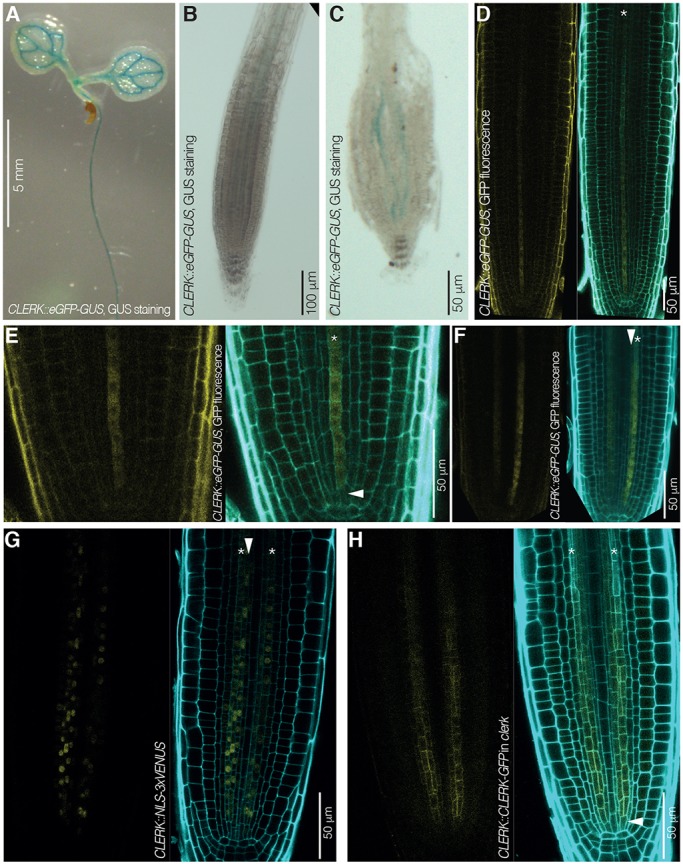
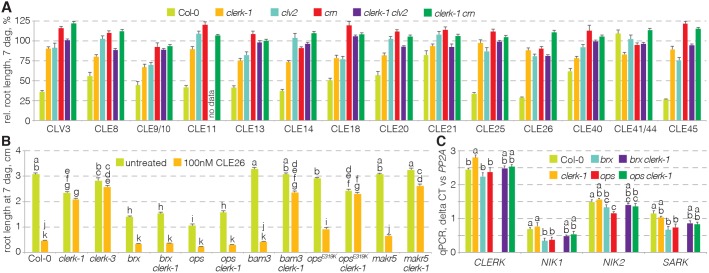
CLAVATA3/EMBRYO SURROUNDING REGION (CLE) peptides are secreted endogenous plant ligands that are sensed by receptor kinases (RKs) to convey environmental and developmental inputs. Typically, this involves an RK with narrow ligand specificity that signals together with a more promiscuous co-receptor. For most CLEs, biologically relevant (co-)receptors are unknown. The dimer of the receptor-like protein CLAVATA 2 (CLV2) and the pseudokinase CORYNE (CRN) conditions perception of so-called root-active CLE peptides, the exogenous application of which suppresses root growth by preventing protophloem formation in the meristem. clv2 as well as crn null mutants are resistant to root-active CLE peptides, possibly because CLV2-CRN promotes expression of their cognate receptors. Here, we have identified the CLE-RESISTANT RECEPTOR KINASE (CLERK) gene, which is required for full sensing of root-active CLE peptides in early developing protophloem. CLERK protein can be replaced by its close homologs, SENESCENCE-ASSOCIATED RECEPTOR-LIKE KINASE (SARK) and NSP-INTERACTING KINASE 1 (NIK1). Yet neither CLERK nor NIK1 ectodomains interact biochemically with described CLE receptor ectodomains. Consistently, CLERK also acts genetically independently of CLV2-CRN We, thus, have discovered a novel hub for redundant CLE sensing in the root.
Keywords: Arabidopsis; BAM3; CLE peptide; Protophloem; Receptor kinase; Root.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
