

Molecular Mechanisms Preventing Senescence in Response to Prolonged Darkness in a Desiccation-Tolerant Plant
- PMID: 29789435
- PMCID: PMC6053018
- DOI: 10.1104/pp.18.00055
Molecular Mechanisms Preventing Senescence in Response to Prolonged Darkness in a Desiccation-Tolerant Plant
Abstract
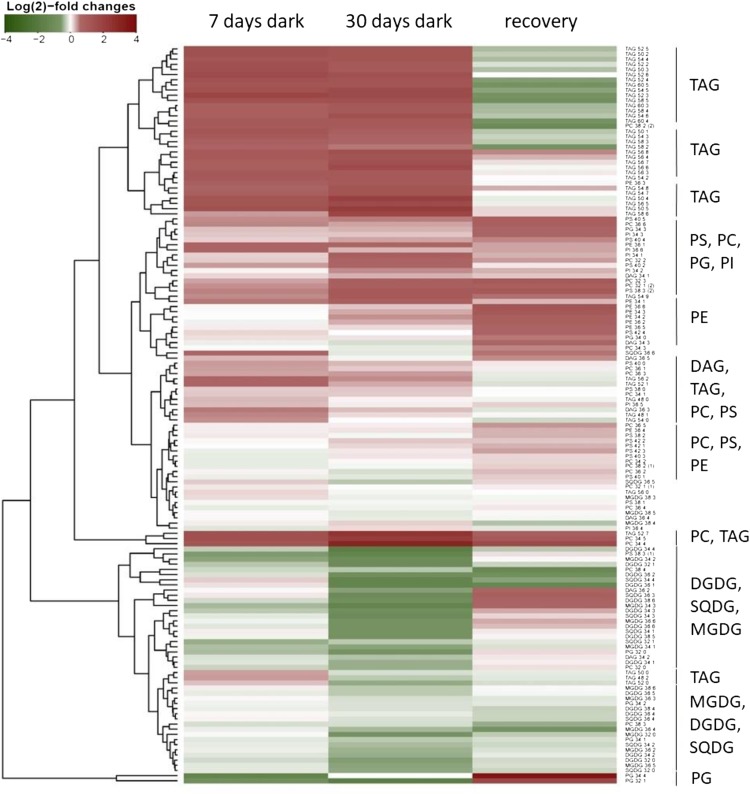
The desiccation-tolerant plant Haberlea rhodopensis can withstand months of darkness without any visible senescence. Here, we investigated the molecular mechanisms of this adaptation to prolonged (30 d) darkness and subsequent return to light. H. rhodopensis plants remained green and viable throughout the dark treatment. Transcriptomic analysis revealed that darkness regulated several transcription factor (TF) genes. Stress- and autophagy-related TFs such as ERF8, HSFA2b, RD26, TGA1, and WRKY33 were up-regulated, while chloroplast- and flowering-related TFs such as ATH1, COL2, COL4, RL1, and PTAC7 were repressed. PHYTOCHROME INTERACTING FACTOR4, a negative regulator of photomorphogenesis and promoter of senescence, also was down-regulated. In response to darkness, most of the photosynthesis- and photorespiratory-related genes were strongly down-regulated, while genes related to autophagy were up-regulated. This occurred concomitant with the induction of SUCROSE NON-FERMENTING1-RELATED PROTEIN KINASES (SnRK1) signaling pathway genes, which regulate responses to stress-induced starvation and autophagy. Most of the genes associated with chlorophyll catabolism, which are induced by darkness in dark-senescing species, were either unregulated (PHEOPHORBIDE A OXYGENASE, PAO; RED CHLOROPHYLL CATABOLITE REDUCTASE, RCCR) or repressed (STAY GREEN-LIKE, PHEOPHYTINASE, and NON-YELLOW COLORING1). Metabolite profiling revealed increases in the levels of many amino acids in darkness, suggesting increased protein degradation. In darkness, levels of the chloroplastic lipids digalactosyldiacylglycerol, monogalactosyldiacylglycerol, phosphatidylglycerol, and sulfoquinovosyldiacylglycerol decreased, while those of storage triacylglycerols increased, suggesting degradation of chloroplast membrane lipids and their conversion to triacylglycerols for use as energy and carbon sources. Collectively, these data show a coordinated response to darkness, including repression of photosynthetic, photorespiratory, flowering, and chlorophyll catabolic genes, induction of autophagy and SnRK1 pathways, and metabolic reconfigurations that enable survival under prolonged darkness.
© 2018 American Society of Plant Biologists. All rights reserved.
Figures









References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215: 403–410 - PubMed
-
- Araújo WL, Ishizaki K, Nunes-Nesi A, Larson TR, Tohge T, Krahnert I, Witt S, Obata T, Schauer N, Graham IA, et al. (2010) Identification of the 2-hydroxyglutarate and isovaleryl-CoA dehydrogenases as alternative electron donors linking lysine catabolism to the electron transport chain of Arabidopsis mitochondria. Plant Cell 22: 1549–1563 - PMC - PubMed
-
- Araújo WL, Ishizaki K, Nunes-Nesi A, Tohge T, Larson TR, Krahnert I, Balbo I, Witt S, Dörmann P, Graham IA, et al. (2011) Analysis of a range of catabolic mutants provides evidence that phytanoyl-coenzyme A does not act as a substrate of the electron-transfer flavoprotein/electron-transfer flavoprotein:ubiquinone oxidoreductase complex in Arabidopsis during dark-induced senescence. Plant Physiol 157: 55–69 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
