

miR172 downregulates the translation of cleistogamy 1 in barley
- PMID: 29790929
- PMCID: PMC6070043
- DOI: 10.1093/aob/mcy058
miR172 downregulates the translation of cleistogamy 1 in barley
Abstract
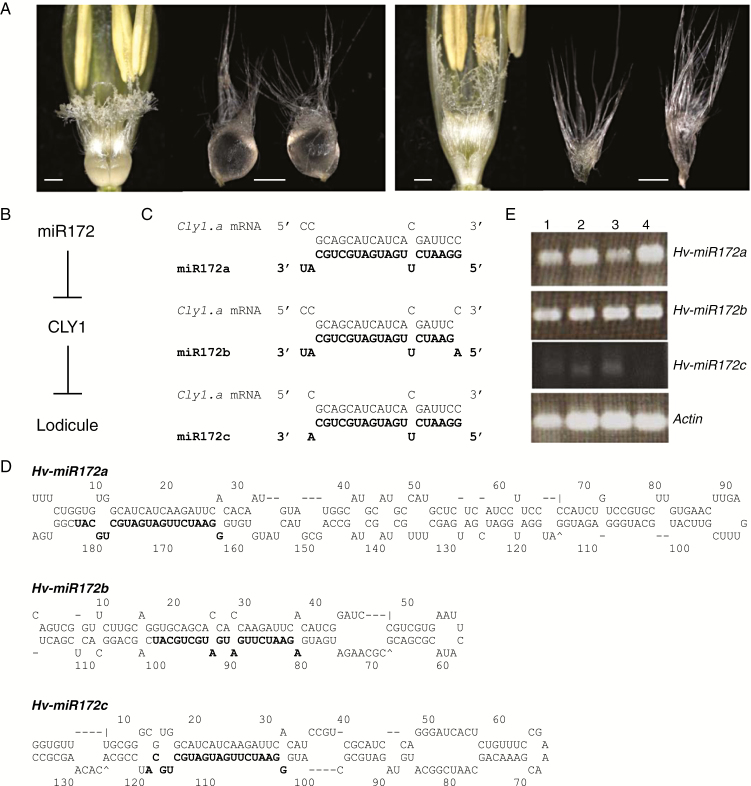
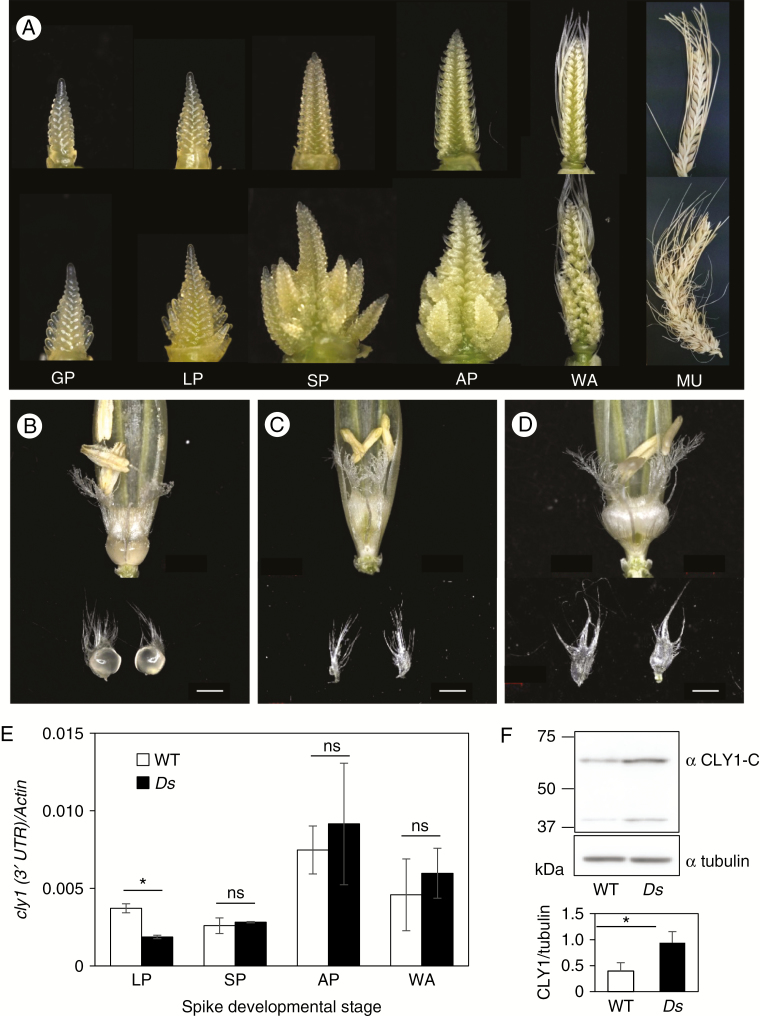
Background and aims: Floret opening in barley is induced by the swelling of the lodicule, a trait under the control of the cleistogamy1 (cly1) gene. The product of cly1 is a member of the APETALA2 (AP2) transcription factor family, which inhibits lodicule development. A sequence polymorphism at the miR172 target site within cly1 has been associated with variation in lodicule development and hence with the cleistogamous phenotype. It was unclear whether miR172 actually functions in cly1 regulation and, if it does, which miR172 gene contributes to cleistogamy. It was also interesting to explore whether miR172-mediated cly1 regulation occurs at transcriptional level or at translational level.
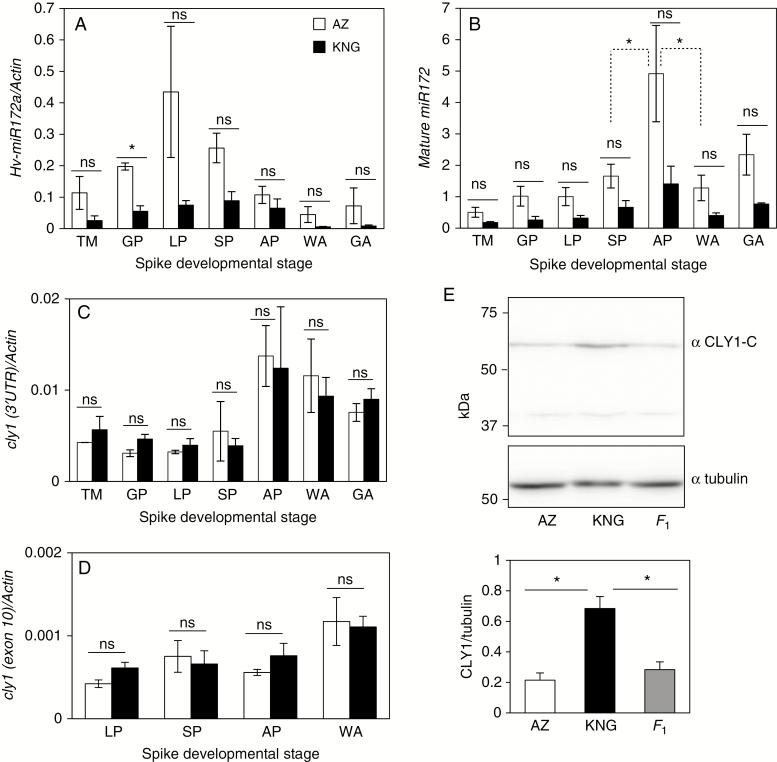
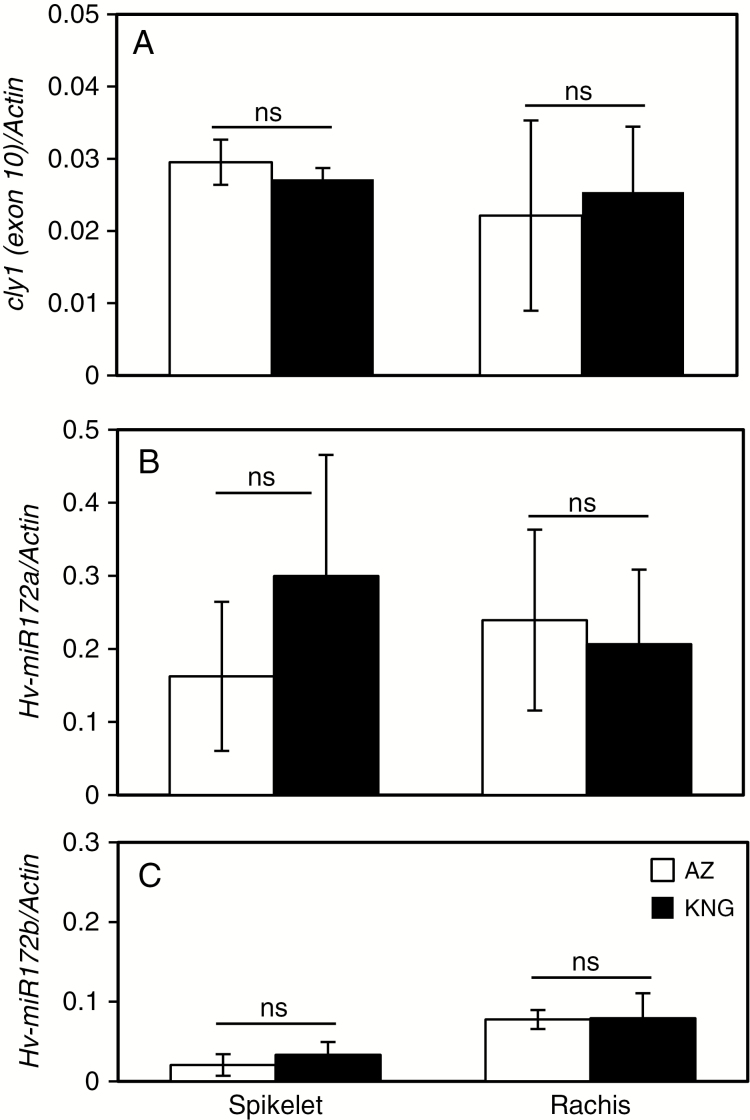
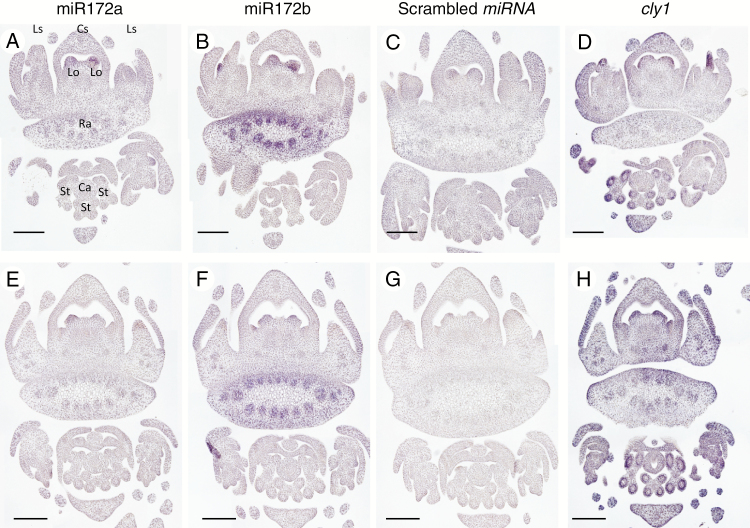
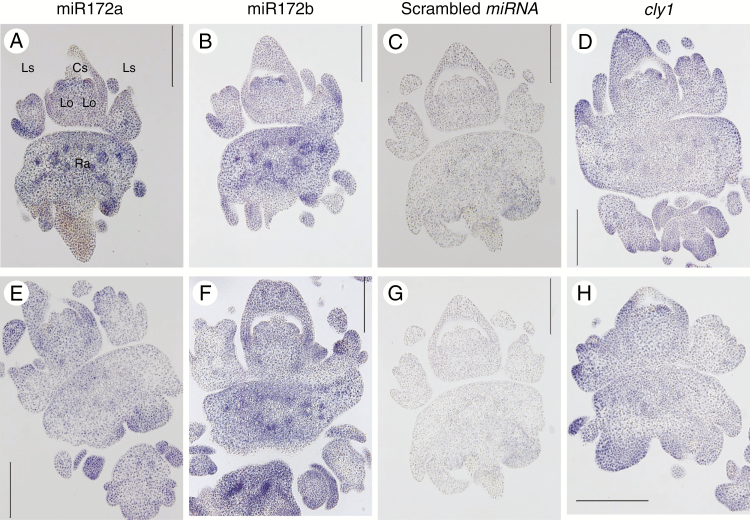
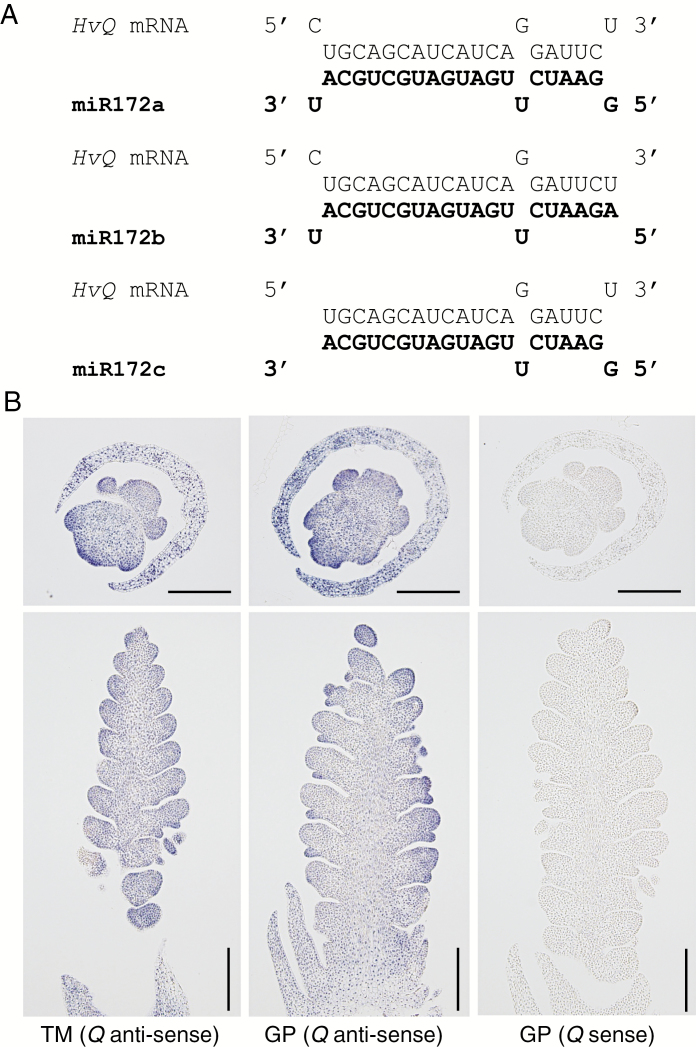
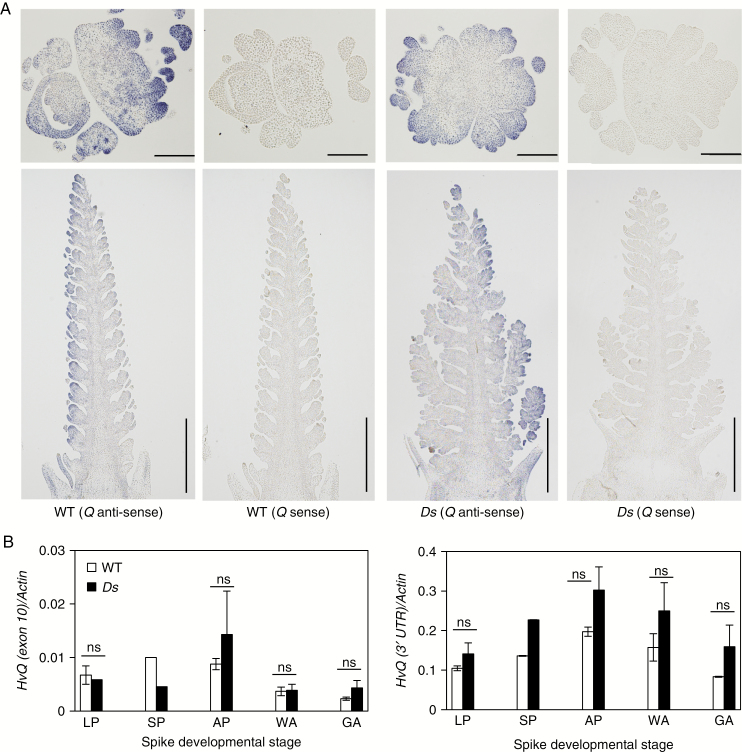
Methods: Deep sequencing of small RNA identified the miR172 sequences expressed in barley immature spikes. miR172 genes were confirmed by computational and expression analysis. miR172 and cly1 expression profiles were determined by in situ hybridization and quantitative expression analysis. Immunoblot analysis provided the CLY1 protein quantifications. Definitive evidence of the role of miR172 in cleistogamy was provided by a transposon Ds-induced mutant of Hv-miR172a.
Key results: A small RNA analysis of the immature barley spike revealed three isomers, miR172a, b and c, of which miR172a was the most abundant. In situ hybridization analysis showed that miR172 and cly1 co-localize in the lodicule primordium, suggesting that these two molecules potentially interact with one another. Immunoblot analysis showed that the sequence polymorphism at the miR172 target site within cly1 reduced the abundance of the CLY1 protein, but not that of its transcript. In a Ds-induced mutant of Hv-miR172a, which generates no mature miR172a, the lodicules fail to grow, resulting in a very small lodicule.
Conclusions: Direct evidence is presented to show that miR172a acts to reduce the abundance of the CLY1 protein, which enables open flowering in barley.
Figures
References
-
- Abdel-Ghani AH, Parzies HK, Omary A, Geiger HH. 2004. Estimating the outcrossing rate of barley landraces and wild barley populations collected from ecologically different regions of Jordan. Theoretical and Applied Genetics 109: 588–595. - PubMed
-
- Ambros V, Lee RC, Lavanway A, Williams PT, Jewell D. 2003. MicroRNAs and other tiny endogenous RNAs in C. elegans. Current Biology 13: 807–818. - PubMed
-
- Bartel DP. 2004. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116: 281–297 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
