

A Central Role for Heme Oxygenase-1 in the Control of Intestinal Epithelial Chemokine Expression
- PMID: 29791903
- PMCID: PMC6008212
- DOI: 10.1159/000488914
A Central Role for Heme Oxygenase-1 in the Control of Intestinal Epithelial Chemokine Expression
Abstract
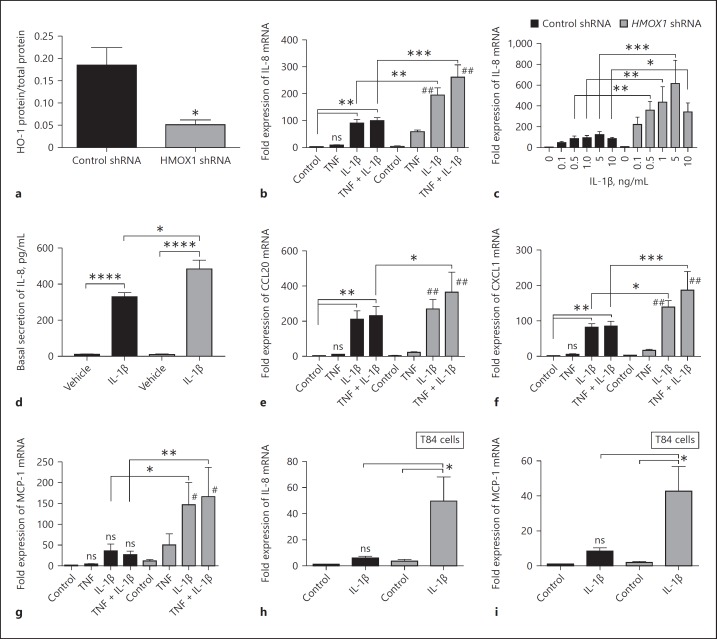
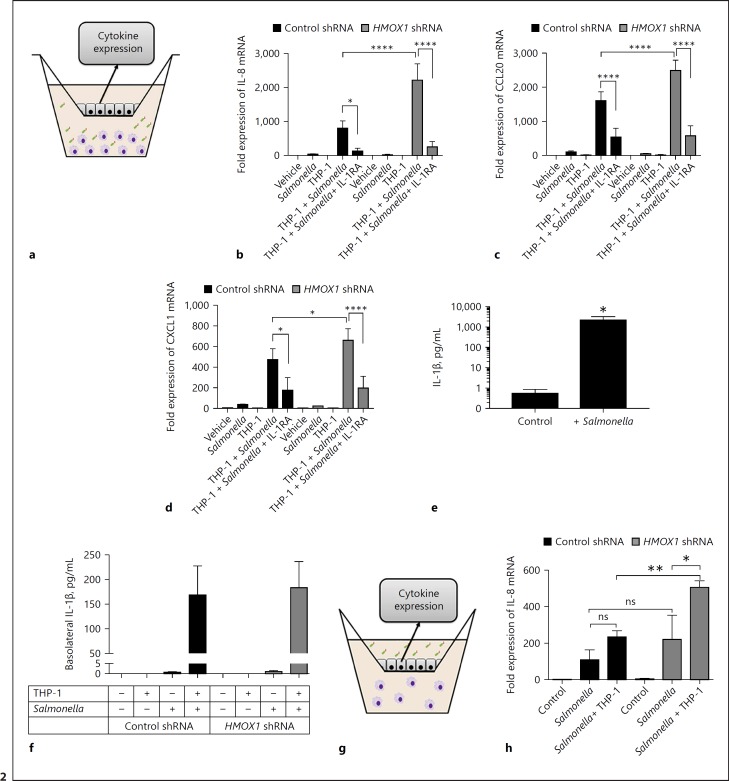
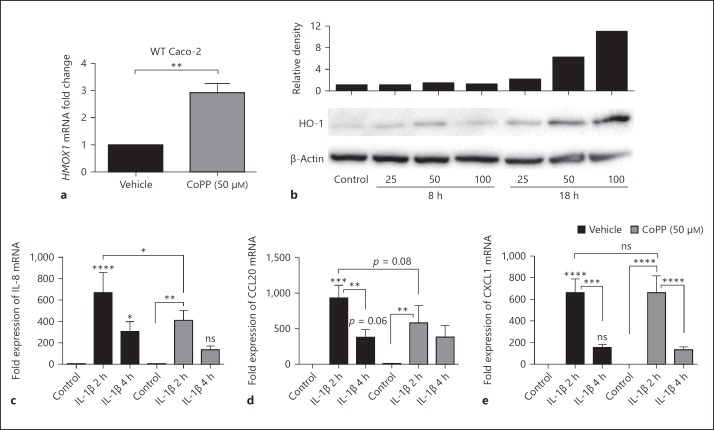
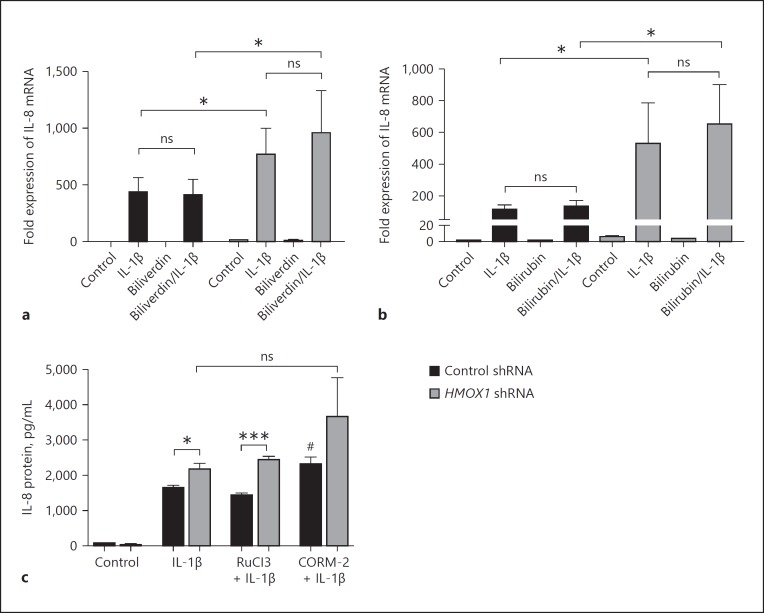
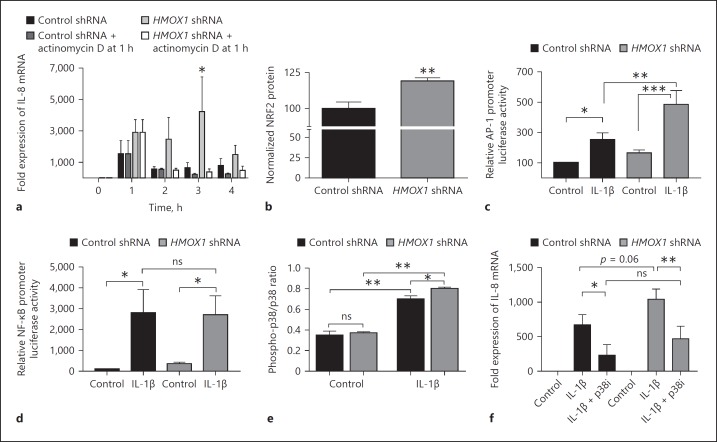
In mucosal inflammatory disorders, the protective influence of heme oxygenase-1 (HO-1) and its metabolic byproducts, carbon monoxide (CO) and biliverdin, is a topic of significant interest. Mechanisms under investigation include the regulation of macrophage function and mucosal cytokine expression. While there is an increasing recognition of the importance of epithelial-derived factors in the maintenance of intestinal mucosal homeostasis, the contribution of intestinal epithelial cell (IEC) HO-1 on inflammatory responses has not previously been investigated. We examined the influence of modulating HO-1 expression on the inflammatory response of human IECs. Engineered deficiency of HO-1 in Caco-2 and T84 IECs led to increased proinflammatory chemokine expression in response to pathogenic bacteria and inflammatory cytokine stimulation. Crosstalk with activated leukocytes also led to increased chemokine expression in HO-1-deficient cells in an IL-1β dependent manner. Treatment of Caco-2 cells with a pharmacological inducer of HO-1 led to the inhibition of chemokine expression. Mechanistic studies suggest that HO-1 and HO-1-related transcription factors, but not HO-1 metabolic products, are partly responsible for the influence of HO-1 on chemokine expression. In conclusion, our data identify HO-1 as a central regulator of IEC chemokine expression that may contribute to homeo-stasis in the intestinal mucosa.
Keywords: Chemokines; Colitis; Cytokines; Homeostasis; Inflammation; Intestinal epithelium.
© 2018 S. Karger AG, Basel.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
