

HIV-1 Tat phosphorylation on Ser-16 residue modulates HIV-1 transcription
- PMID: 29792216
- PMCID: PMC5966876
- DOI: 10.1186/s12977-018-0422-5
HIV-1 Tat phosphorylation on Ser-16 residue modulates HIV-1 transcription
Abstract
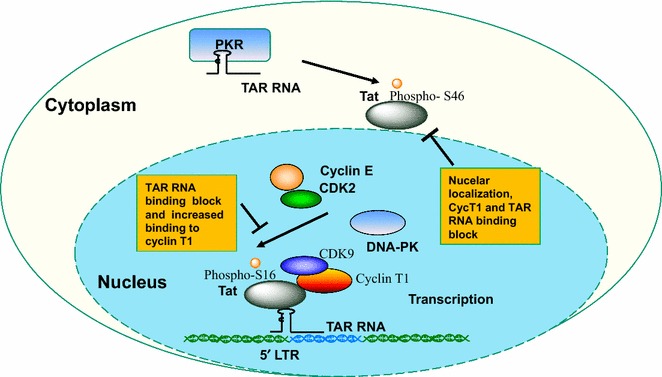
Background: HIV-1 transcription activator protein Tat is phosphorylated in vitro by CDK2 and DNA-PK on Ser-16 residue and by PKR on Tat Ser-46 residue. Here we analyzed Tat phosphorylation in cultured cells and its functionality.
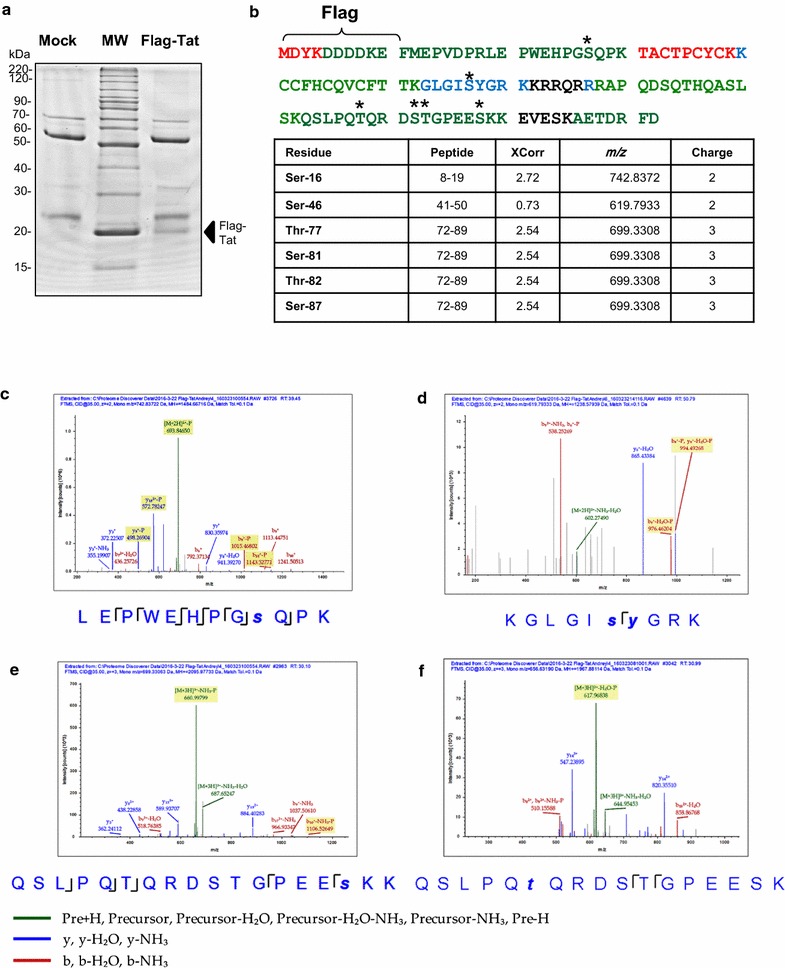
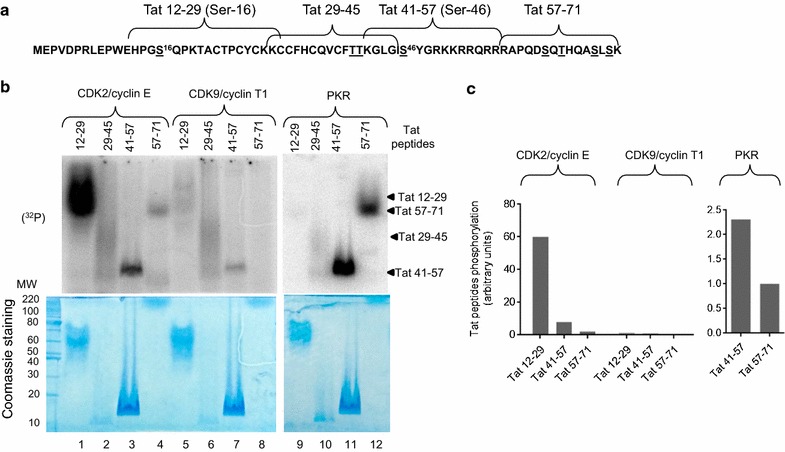
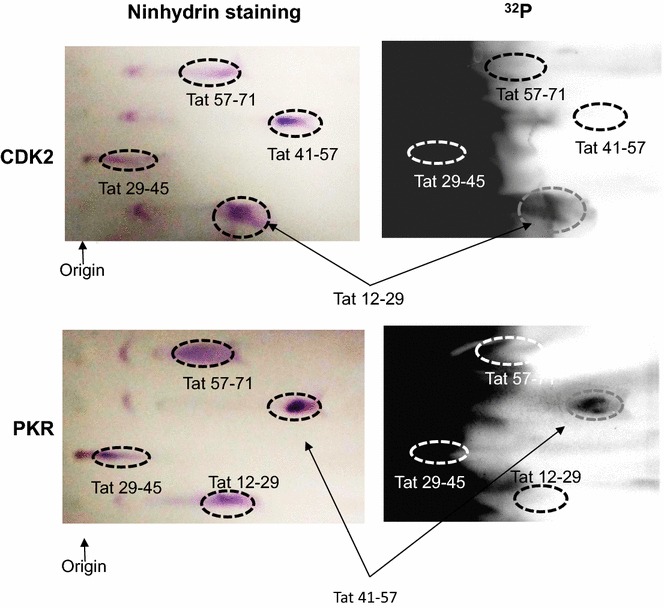
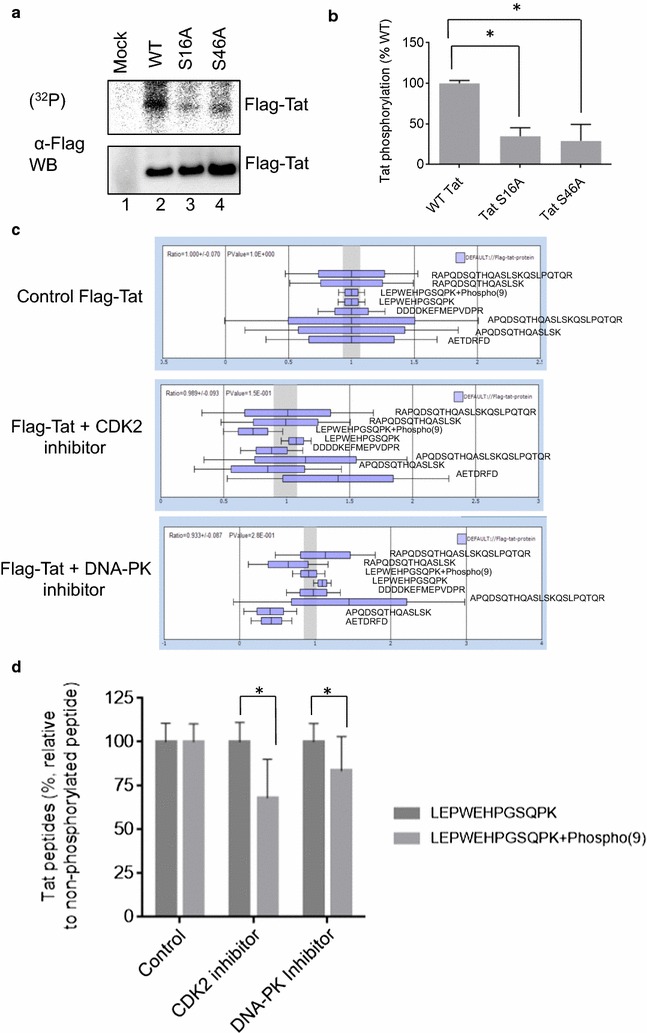
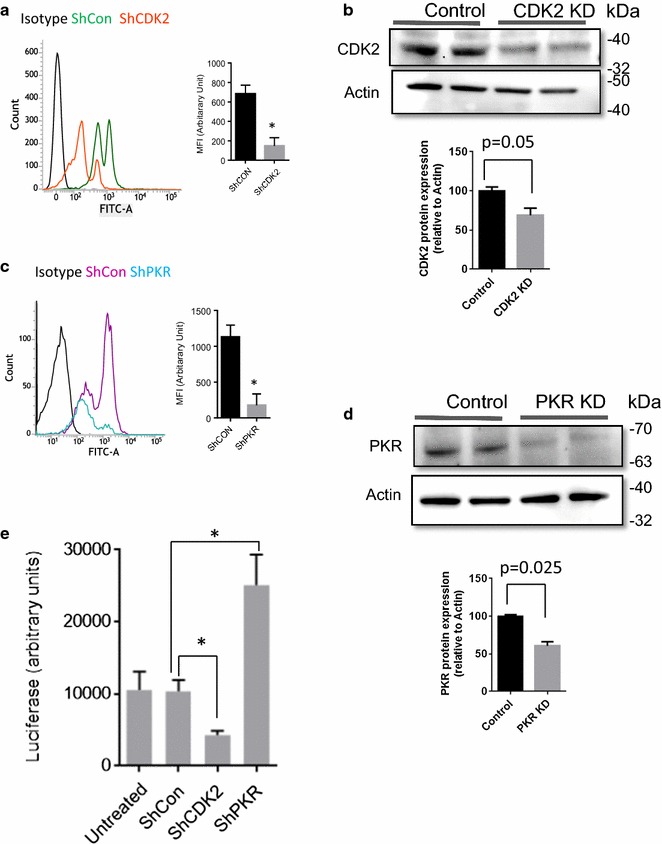
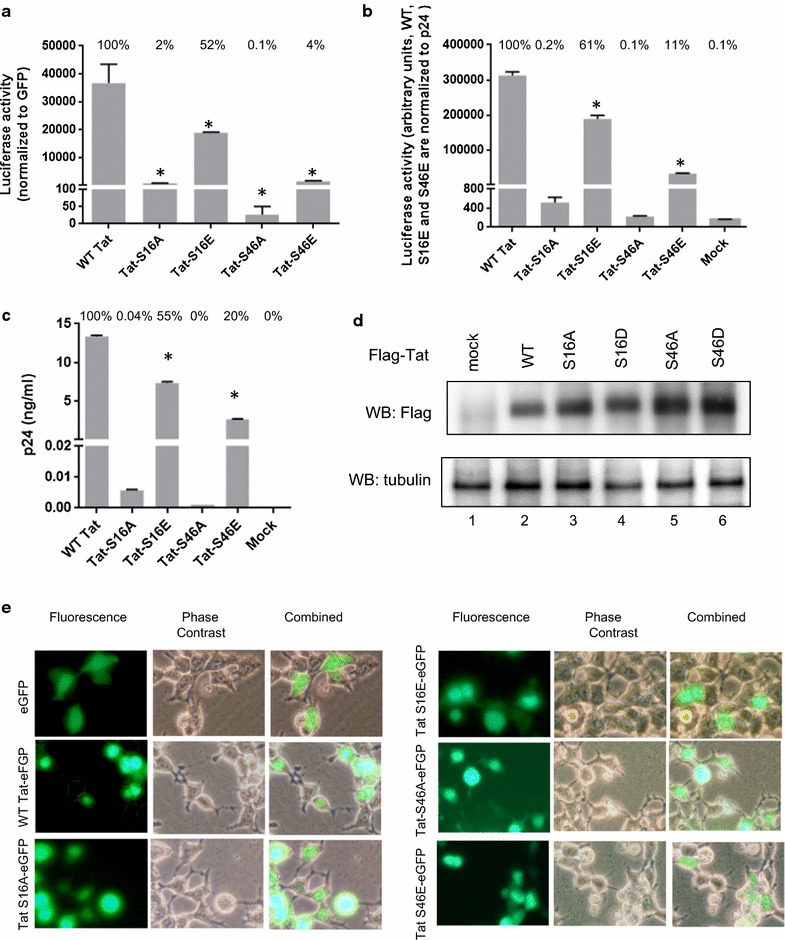
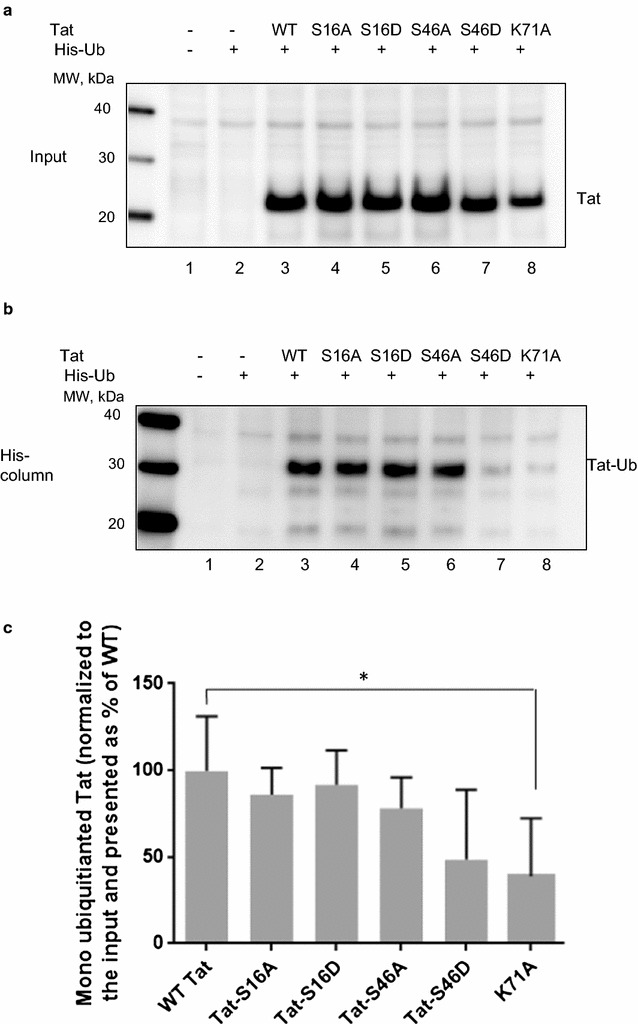
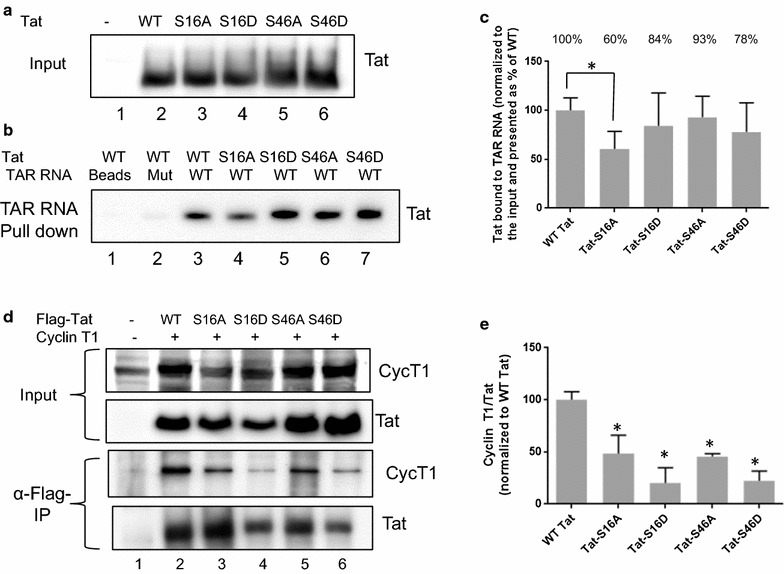
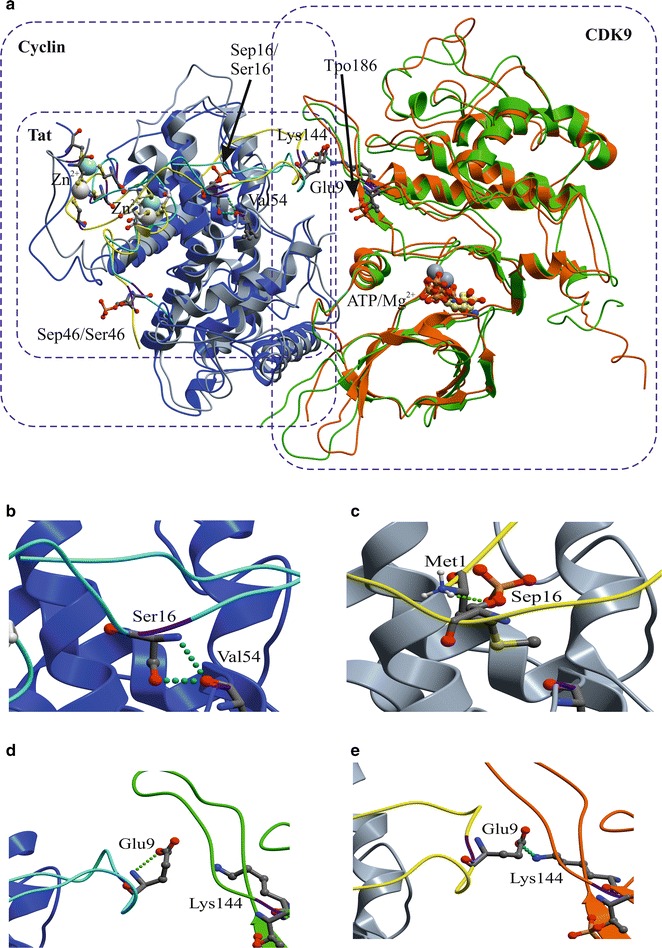
Results: Mass spectrometry analysis showed primarily Tat Ser-16 phosphorylation in cultured cells. In vitro, CDK2/cyclin E predominantly phosphorylated Tat Ser-16 and PKR-Tat Ser-46. Alanine mutations of either Ser-16 or Ser-46 decreased overall Tat phosphorylation. Phosphorylation of Tat Ser-16 was reduced in cultured cells treated by a small molecule inhibitor of CDK2 and, to a lesser extent, an inhibitor of DNA-PK. Conditional knock-downs of CDK2 and PKR inhibited and induced one round HIV-1 replication respectively. HIV-1 proviral transcription was inhibited by Tat alanine mutants and partially restored by S16E mutation. Pseudotyped HIV-1 with Tat S16E mutation replicated well, and HIV-1 Tat S46E-poorly, but no live viruses were obtained with Tat S16A or Tat S46A mutations. TAR RNA binding was affected by Tat Ser-16 alanine mutation. Binding to cyclin T1 showed decreased binding of all Ser-16 and Ser-46 Tat mutants with S16D and Tat S46D mutationts showing the strongest effect. Molecular modelling and molecular dynamic analysis revealed significant structural changes in Tat/CDK9/cyclin T1 complex with phosphorylated Ser-16 residue, but not with phosphorylated Ser-46 residue.
Conclusion: Phosphorylation of Tat Ser-16 induces HIV-1 transcription, facilitates binding to TAR RNA and rearranges CDK9/cyclin T1/Tat complex. Thus, phosphorylation of Tat Ser-16 regulates HIV-1 transcription and may serve as target for HIV-1 therapeutics.
Figures
References
-
- Lafeuillade A, Stevenson M. The search for a cure for persistent HIV reservoirs. AIDS Rev. 2011;13(2):63–66. - PubMed
-
- Deng L, Ammosova T, Pumfery A, Kashanchi F, Nekhai S. HIV-1 Tat interaction with RNA polymerase II C-terminal domain (CTD) and a dynamic association with CDK2 induce CTD phosphorylation and transcription from HIV-1 promoter. J Biol Chem. 2002;277(37):33922–33929. doi: 10.1074/jbc.M111349200. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL125005/HL/NHLBI NIH HHS/United States
- AI117970/National Institute of Allergy and Infectious Diseases/International
- G12 MD007597/MD/NIMHD NIH HHS/United States
- P50 HL118006/HL/NHLBI NIH HHS/United States
- P50HL118006/HL/NHLBI NIH HHS/United States
- U19AI109664/National Institute of Allergy and Infectious Diseases/International
- P30 AI117970/AI/NIAID NIH HHS/United States
- R01HL125005/HL/NHLBI NIH HHS/United States
- 5G12MD007597/MD/NIMHD NIH HHS/United States
- 12-04-91444-NIZ/Russian Foundation for Basic Research/International
- 2012-1.5-12-000-1001-030/Ministry of Education and Science of the Russian Federation/International
- U19 AI109664/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
