

Nsp1α of Porcine Reproductive and Respiratory Syndrome Virus Strain BB0907 Impairs the Function of Monocyte-Derived Dendritic Cells via the Release of Soluble CD83
- PMID: 29793955
- PMCID: PMC6052304
- DOI: 10.1128/JVI.00366-18
Nsp1α of Porcine Reproductive and Respiratory Syndrome Virus Strain BB0907 Impairs the Function of Monocyte-Derived Dendritic Cells via the Release of Soluble CD83
Abstract
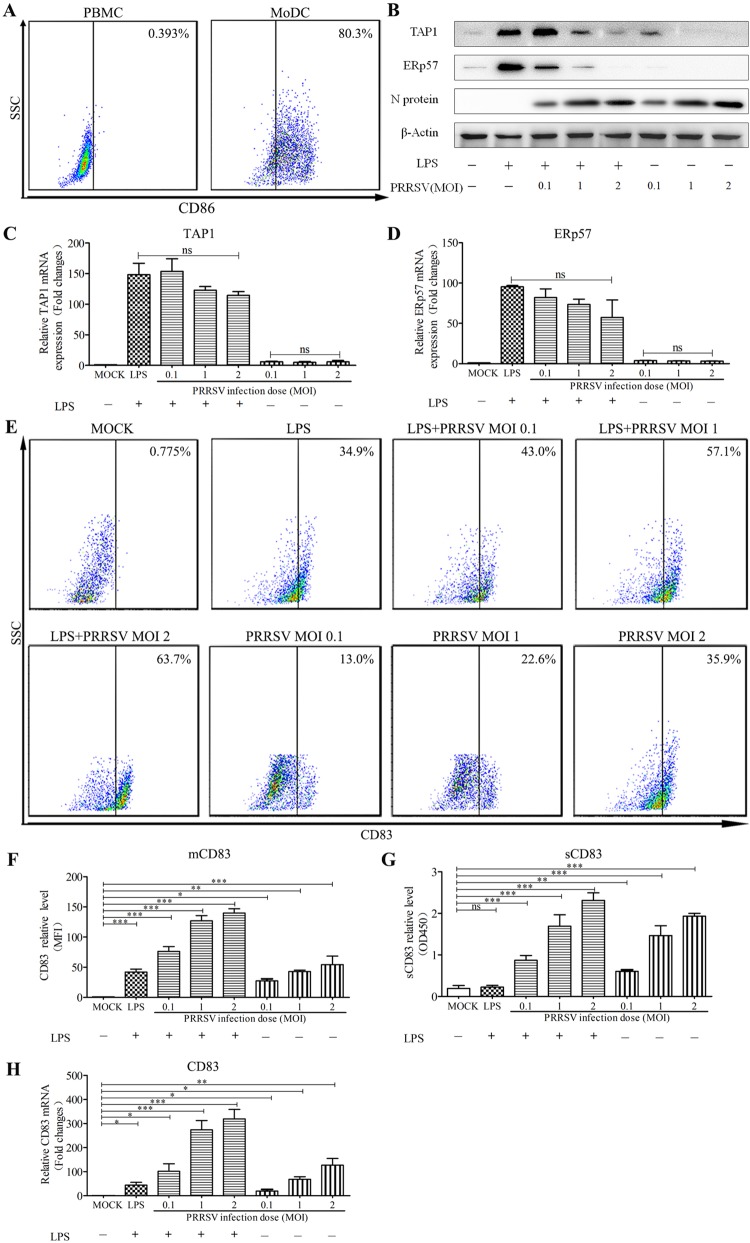
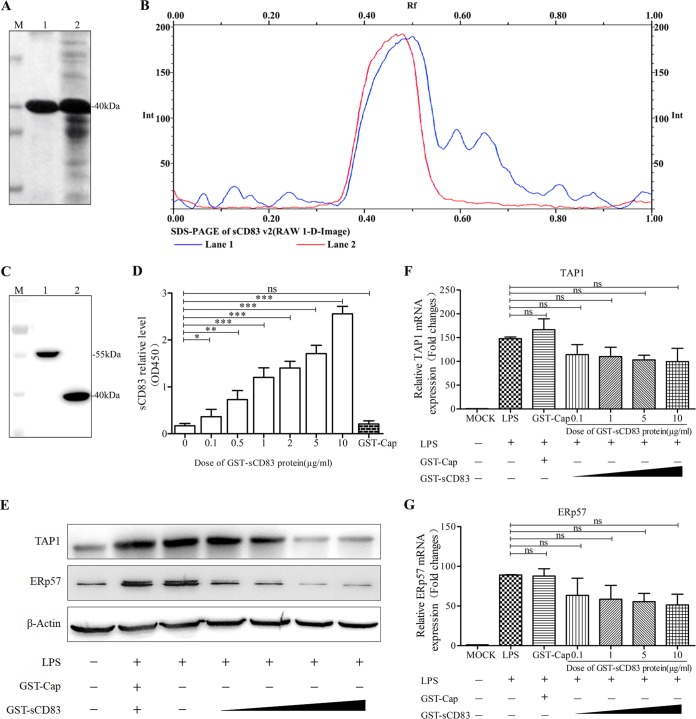
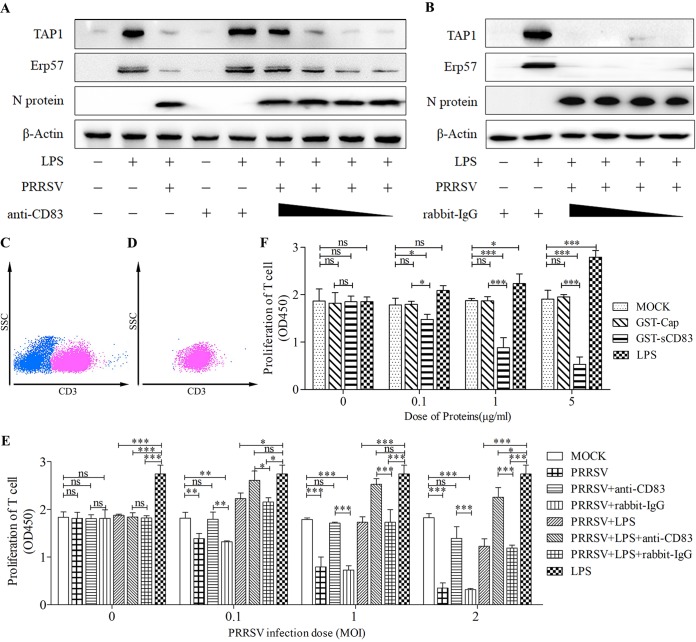
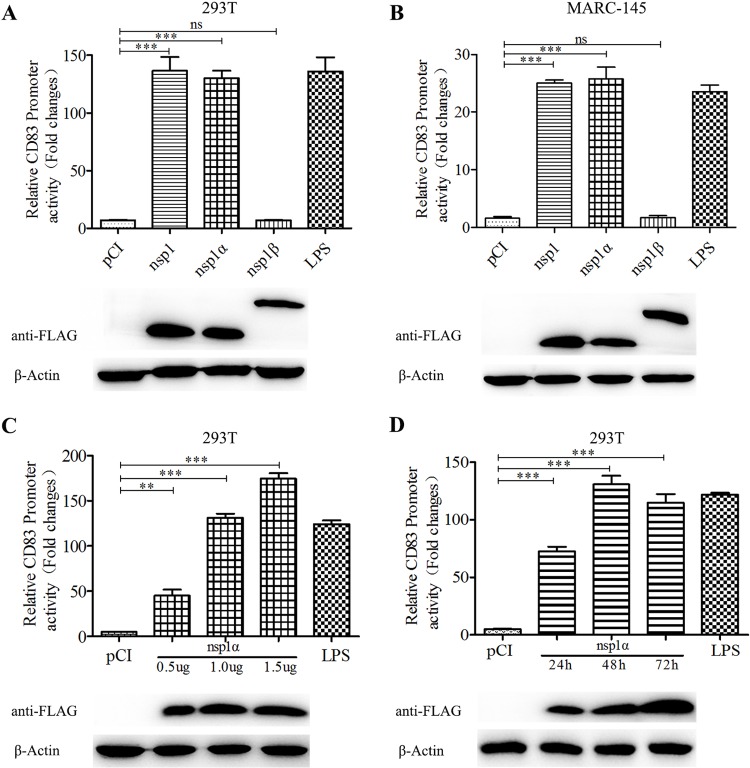
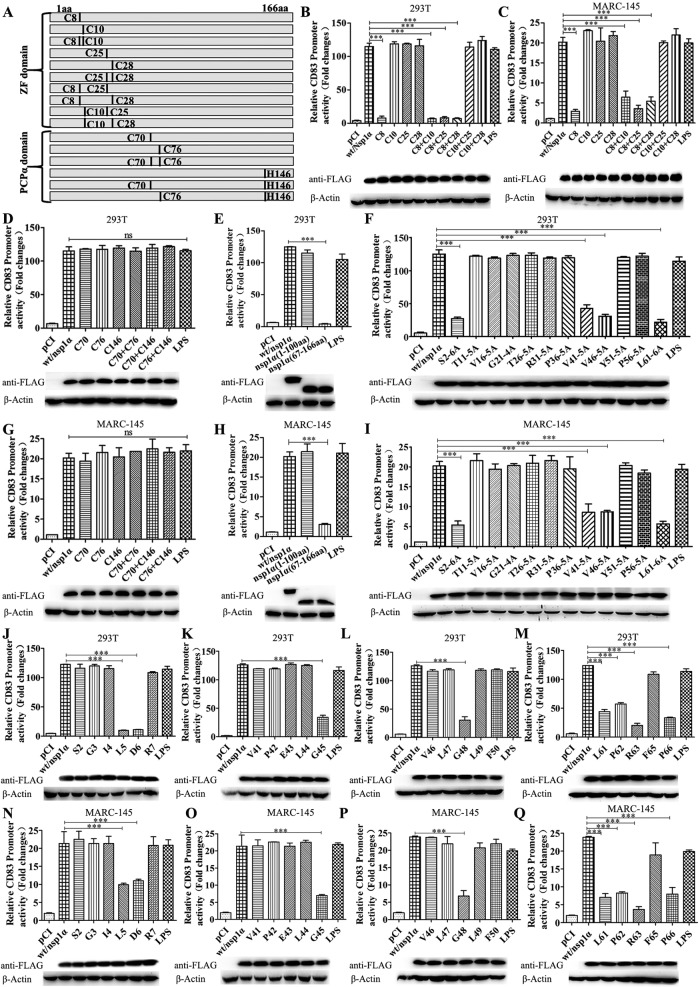
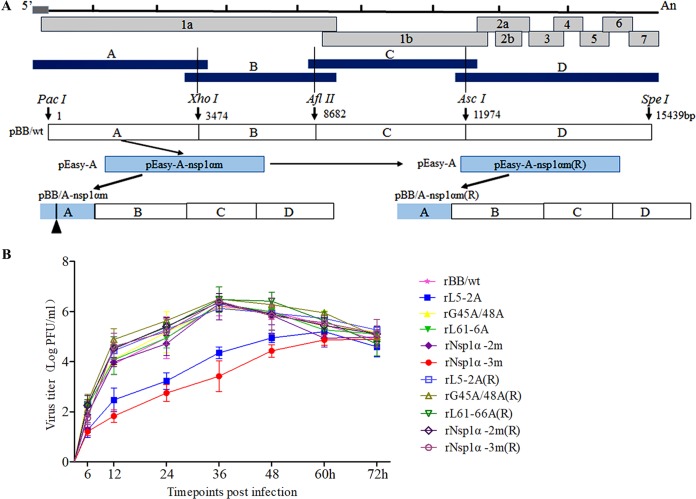
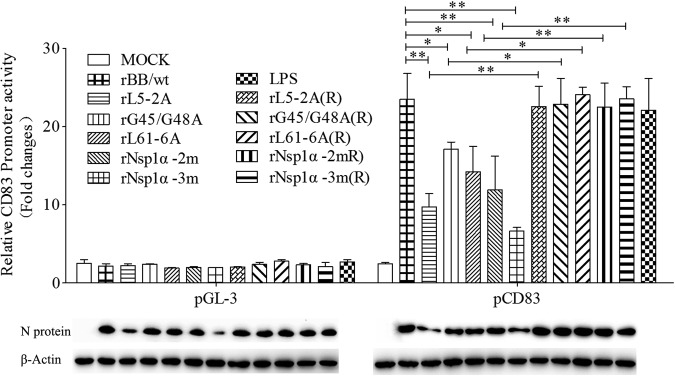
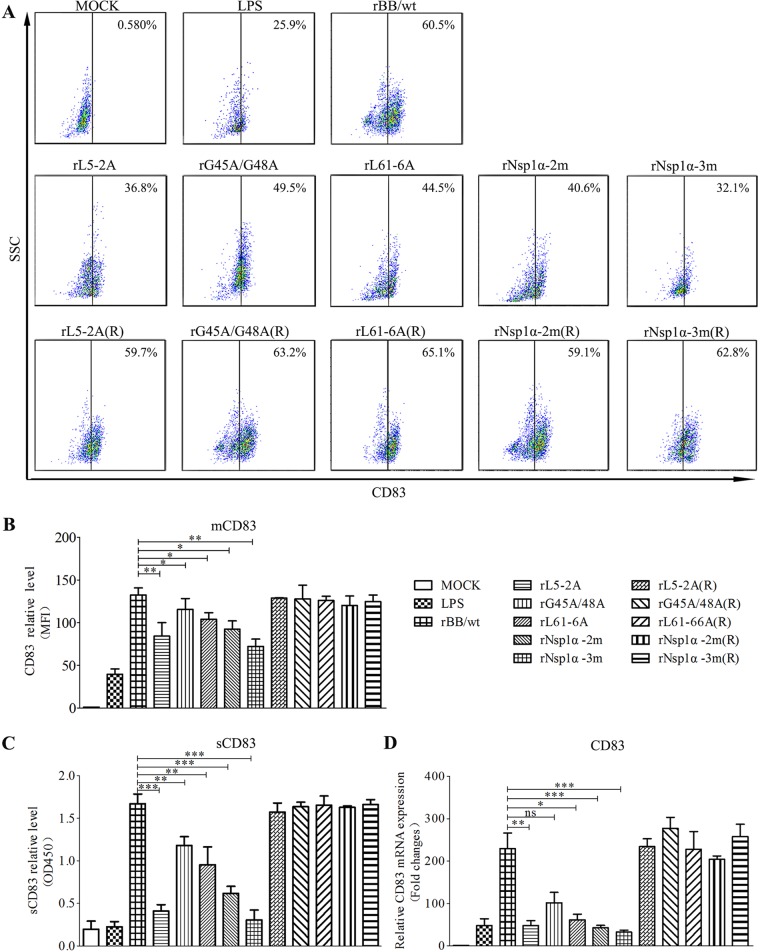
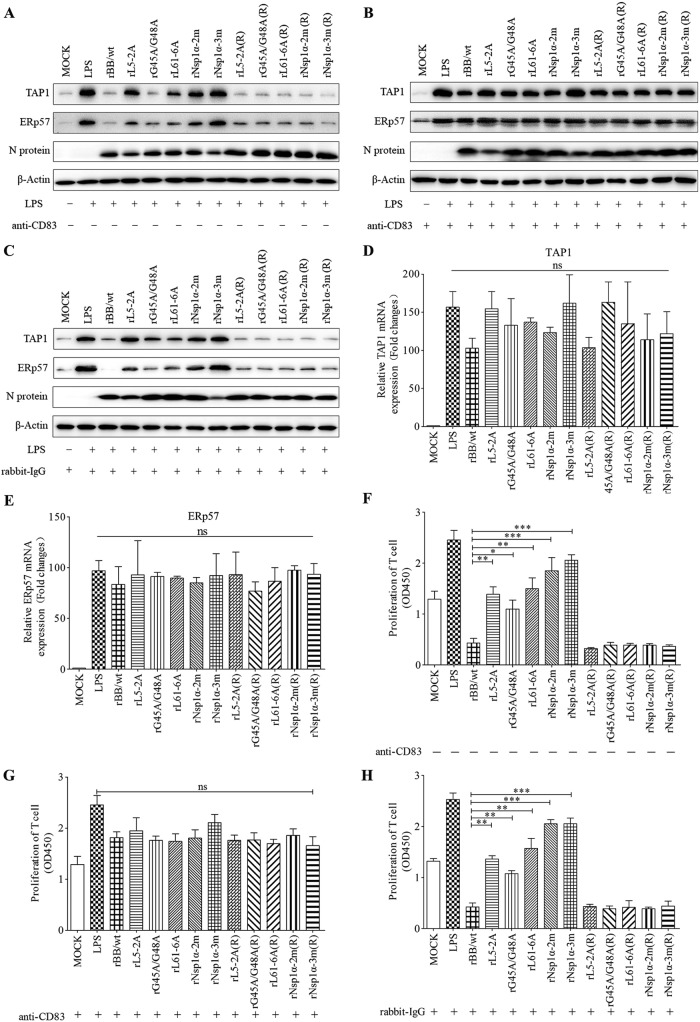
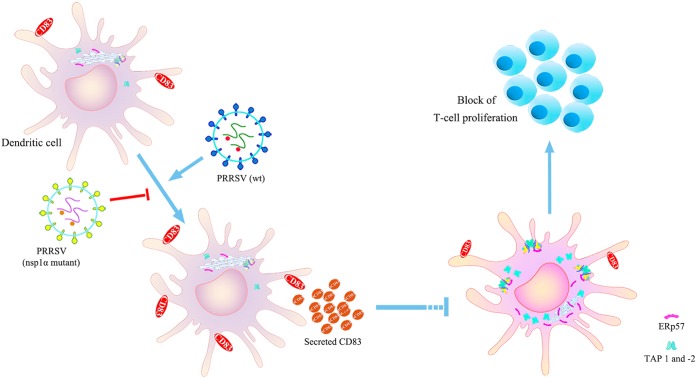
Porcine reproductive and respiratory syndrome virus (PRRSV), a virulent pathogen of swine, suppresses the innate immune response and induces persistent infection. One mechanism used by viruses to evade the immune system is to cripple the antigen-processing machinery in monocyte-derived dendritic cells (MoDCs). In this study, we show that MoDCs infected by PRRSV express lower levels of the major histocompatibility complex (MHC)-peptide complex proteins TAP1 and ERp57 and are impaired in their ability to stimulate T cell proliferation and increase their production of CD83. Neutralization of sCD83 removes the inhibitory effects of PRRSV on MoDCs. When MoDCs are incubated with exogenously added sCD83 protein, TAP1 and ERp57 expression decreases and T lymphocyte activation is impaired. PRRSV nonstructural protein 1α (Nsp1α) enhances CD83 promoter activity. Mutations in the ZF domain of Nsp1α abolish its ability to activate the CD83 promoter. We generated recombinant PRRSVs with mutations in Nsp1α and the corresponding repaired PRRSVs. Viruses with Nsp1α mutations did not decrease levels of TAP1 and ERp57, impair the ability of MoDCs to stimulate T cell proliferation, or increase levels of sCD83. We show that the ZF domain of Nsp1α stimulates the secretion of CD83, which in turn inhibits MoDC function. Our study provides new insights into the mechanisms of immune suppression by PRRSV.IMPORTANCE PRRSV has a severe impact on the swine industry throughout the world. Understanding the mechanisms by which PRRSV infection suppresses the immune system is essential for a robust and sustainable swine industry. Here, we demonstrated that PRRSV infection manipulates MoDCs by interfering with their ability to produce proteins in the MHC-peptide complex. The virus also impairs the ability of MoDCs to stimulate cell proliferation, due in large part to the enhanced release of soluble CD83 from PRRSV-infected MoDCs. The viral nonstructural protein 1 (Nsp1) is responsible for upregulating CD83 promoter activity. Amino acids in the ZF domain of Nsp1α (L5-2A, rG45A, G48A, and L61-6A) are essential for CD83 promoter activation. Viruses with mutations at these sites no longer inhibit MoDC-mediated T cell proliferation. These findings provide novel insights into the mechanism by which the adaptive immune response is suppressed during PRRSV infection.
Keywords: Erp57; MoDC; Nsp1α; TAP1; porcine reproductive and respiratory syndrome virus; sCD83.
Copyright © 2018 American Society for Microbiology.
Figures
References
-
- Subramaniam S, Piñeyro P, Derscheid RJ, Madson DM, Magstadt DR, Meng X. 2017. Dendritic cell-targeted porcine reproductive and respiratory syndrome virus (PRRSV) antigens adjuvanted with polyinosinic-polycytidylic acid (poly (I:C)) induced non-protective immune responses against heterologous type 2 PRRSV challenge in pigs. Vet Immunol Immunopathol 190:18–25. doi: 10.1016/j.vetimm.2017.07.003. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
