

Microtubules grow by the addition of bent guanosine triphosphate tubulin to the tips of curved protofilaments
- PMID: 29794031
- PMCID: PMC6080942
- DOI: 10.1083/jcb.201802138
Microtubules grow by the addition of bent guanosine triphosphate tubulin to the tips of curved protofilaments
Abstract
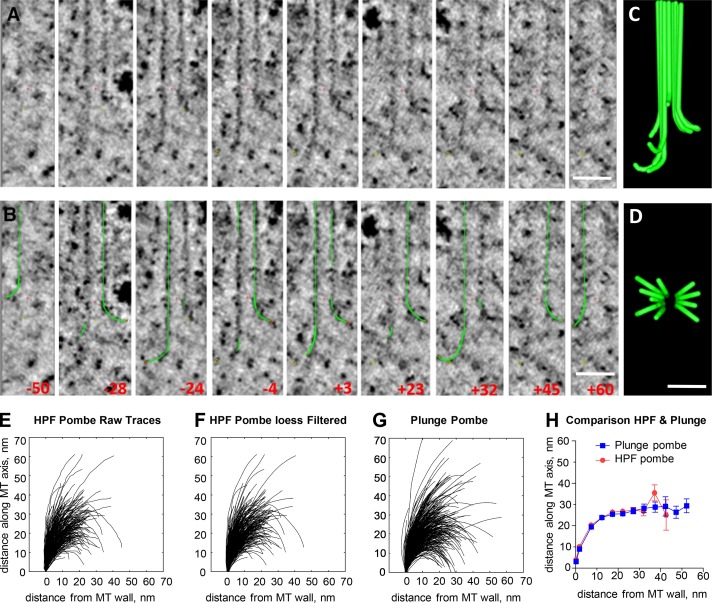
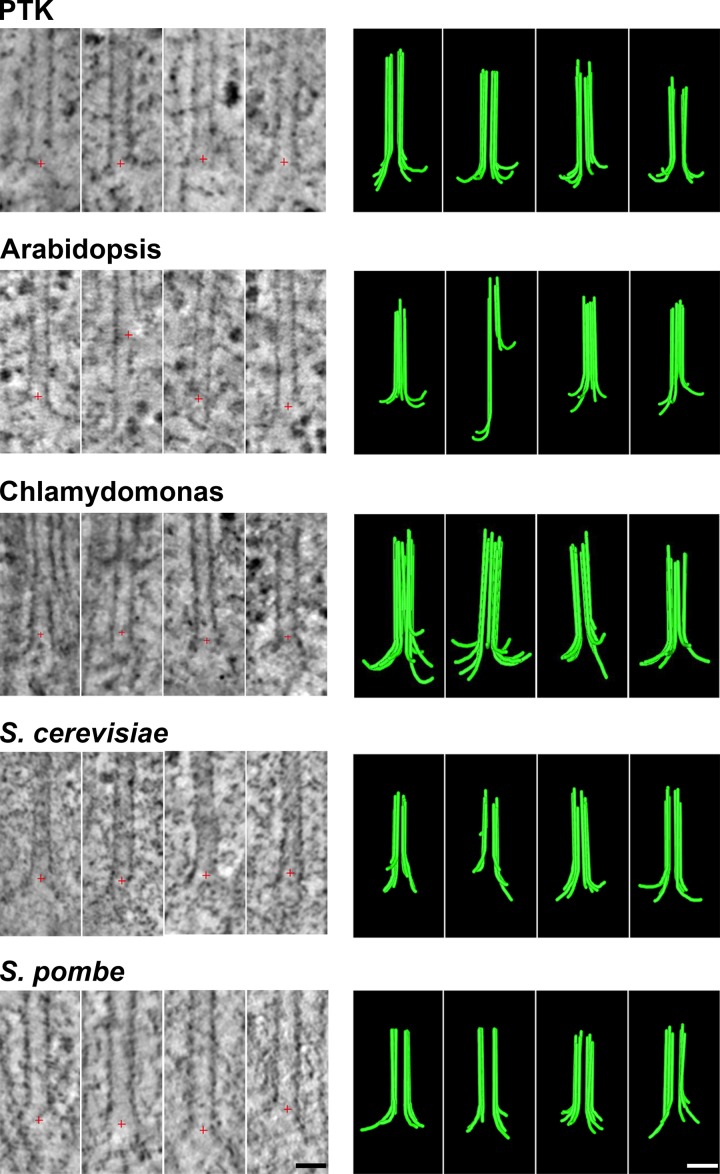
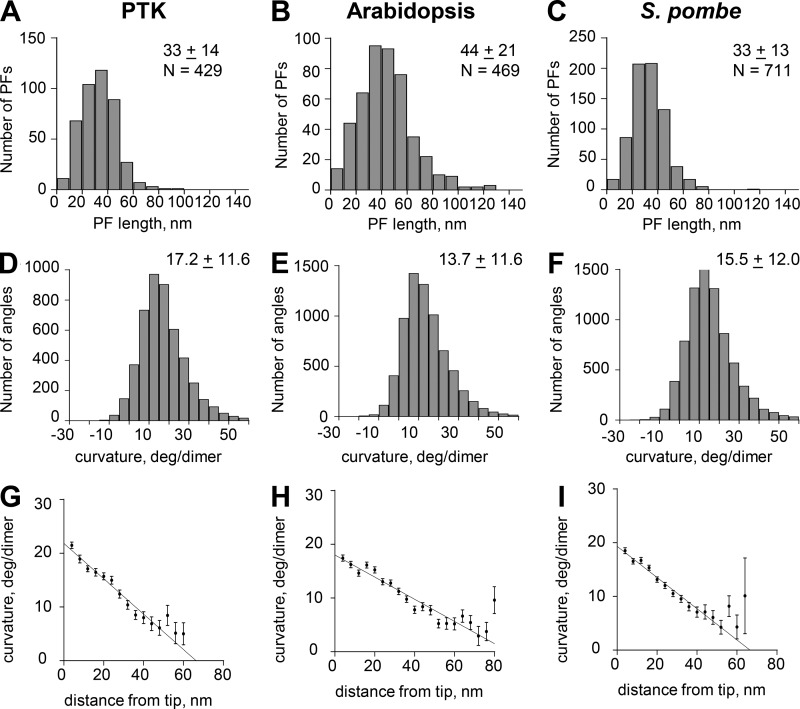
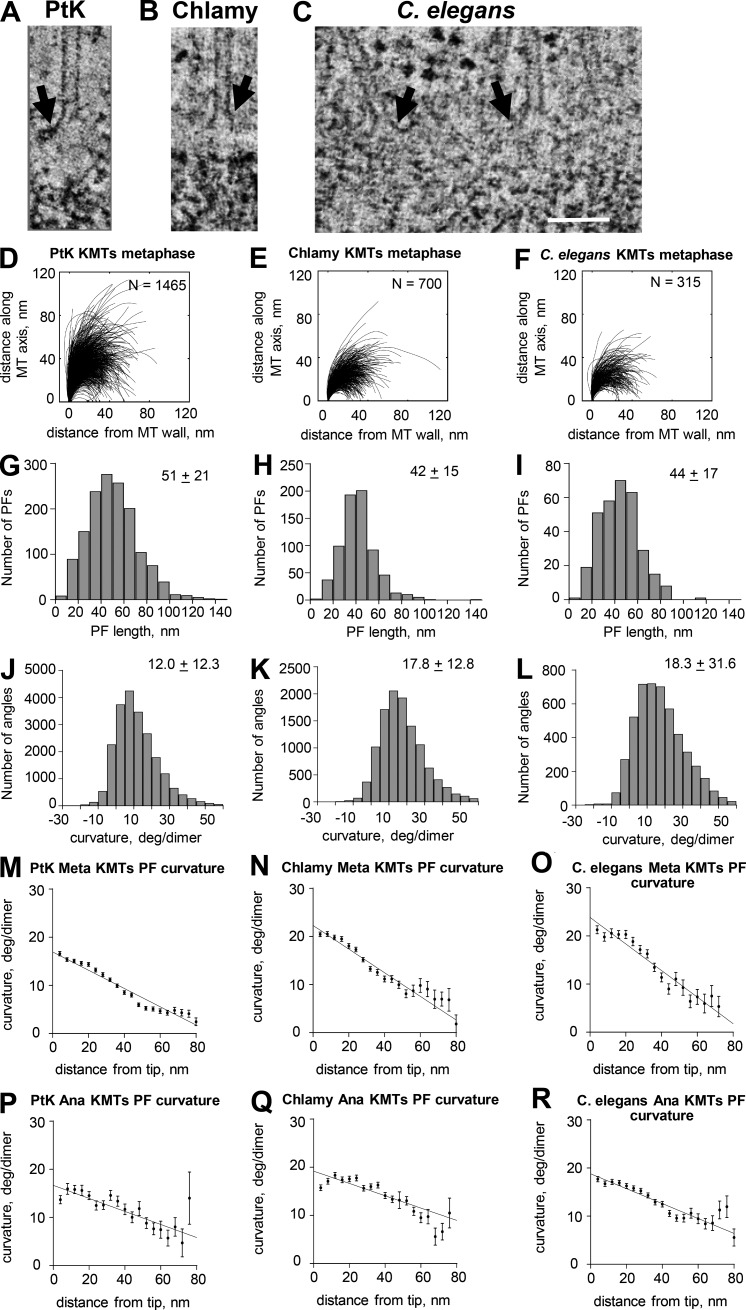
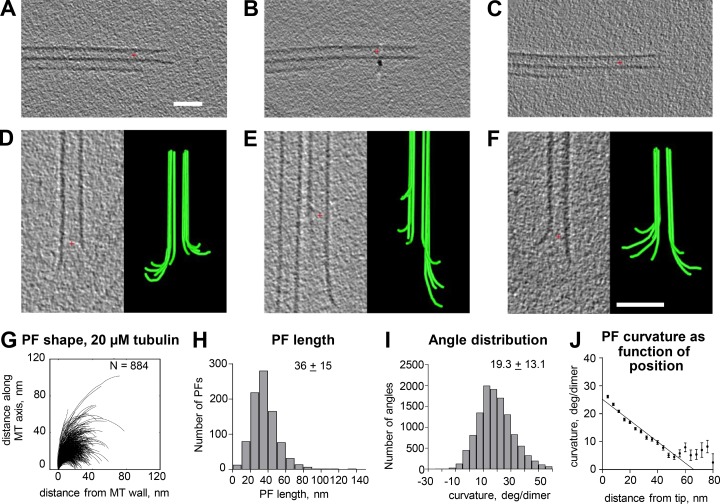
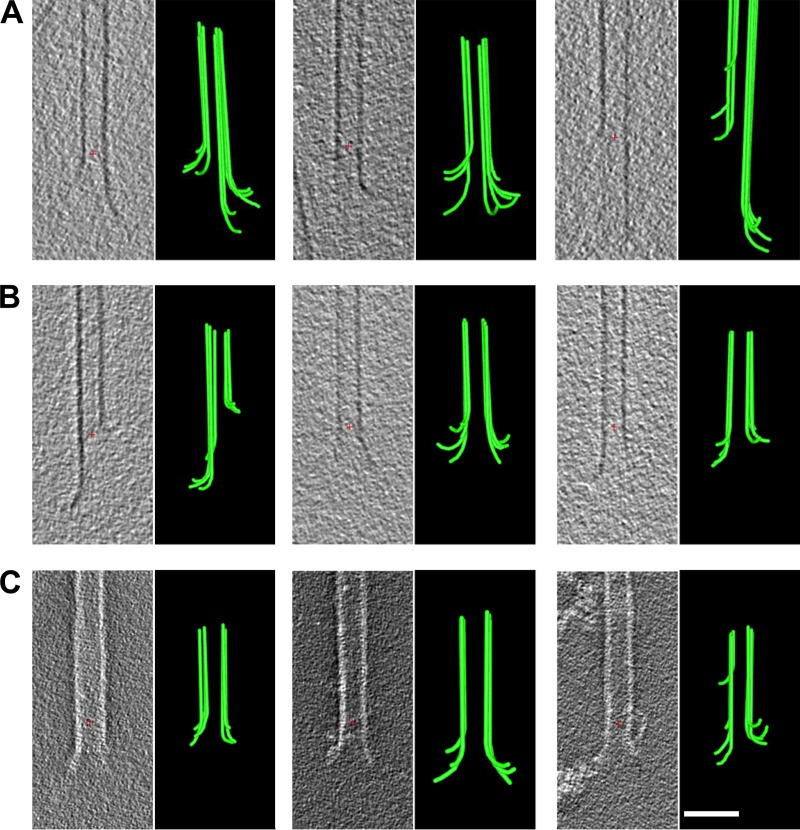
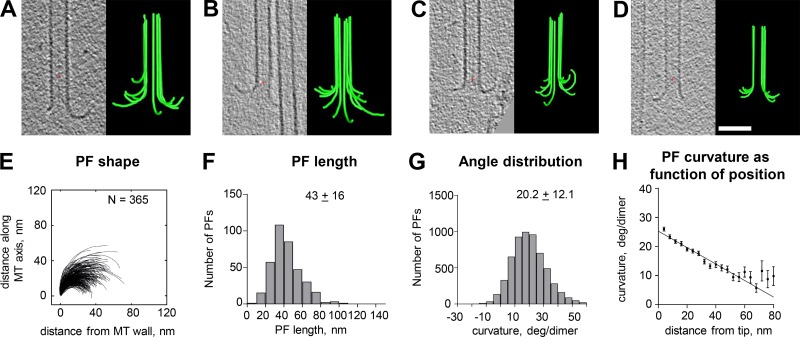
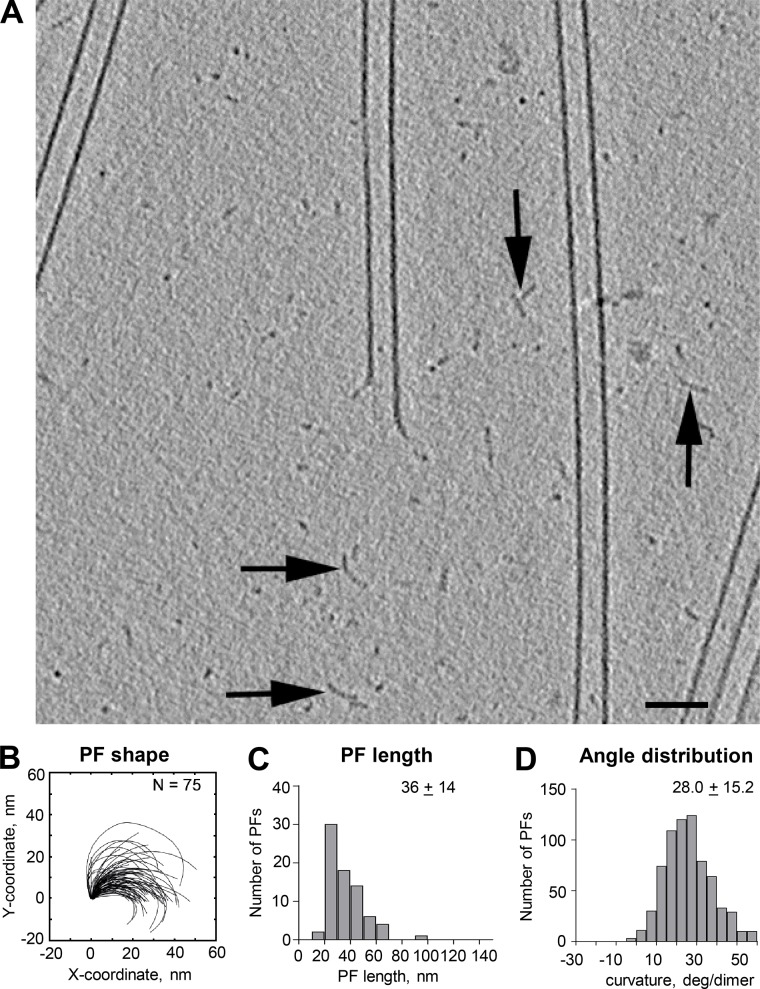
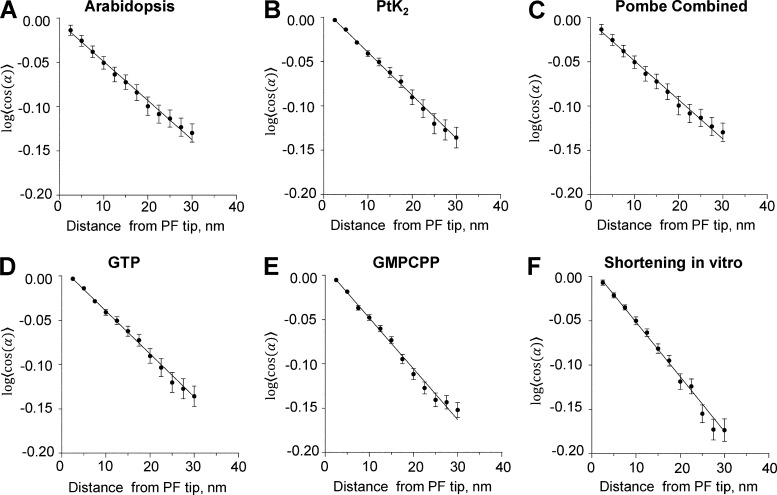
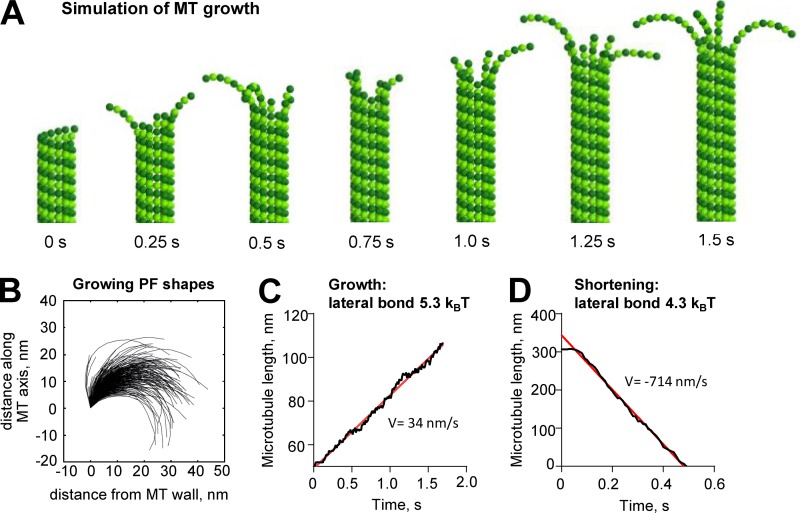
We used electron tomography to examine microtubules (MTs) growing from pure tubulin in vitro as well as two classes of MTs growing in cells from six species. The tips of all these growing MTs display bent protofilaments (PFs) that curve away from the MT axis, in contrast with previously reported MTs growing in vitro whose tips are either blunt or sheetlike. Neither high pressure nor freezing is responsible for the PF curvatures we see. The curvatures of PFs on growing and shortening MTs are similar; all are most curved at their tips, suggesting that guanosine triphosphate-tubulin in solution is bent and must straighten to be incorporated into the MT wall. Variations in curvature suggest that PFs are flexible in their plane of bending but rigid to bending out of that plane. Modeling by Brownian dynamics suggests that PF straightening for MT growth can be achieved by thermal motions, providing a simple mechanism with which to understand tubulin polymerization.
© 2018 McIntosh et al.
Figures
Comment in
-
A new look for the growing microtubule end?J Cell Biol. 2018 Aug 6;217(8):2609-2611. doi: 10.1083/jcb.201807036. Epub 2018 Jul 13. J Cell Biol. 2018. PMID: 30006463 Free PMC article.
References
-
- Atherton J., Jiang K., Stangier M.M., Luo Y., Hua S., Houben K., van Hooff J.J.E., Joseph A.-P., Scarabelli G., Grant B.J., et al. 2017. A structural model for microtubule minus-end recognition and protection by CAMSAP proteins. Nat. Struct. Mol. Biol. 24:931–943. 10.1038/nsmb.3483 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
