

Insights into catalysis and function of phosphoribosyl-linked serine ubiquitination
- PMID: 29795347
- PMCID: PMC5980784
- DOI: 10.1038/s41586-018-0145-8
Insights into catalysis and function of phosphoribosyl-linked serine ubiquitination
Abstract
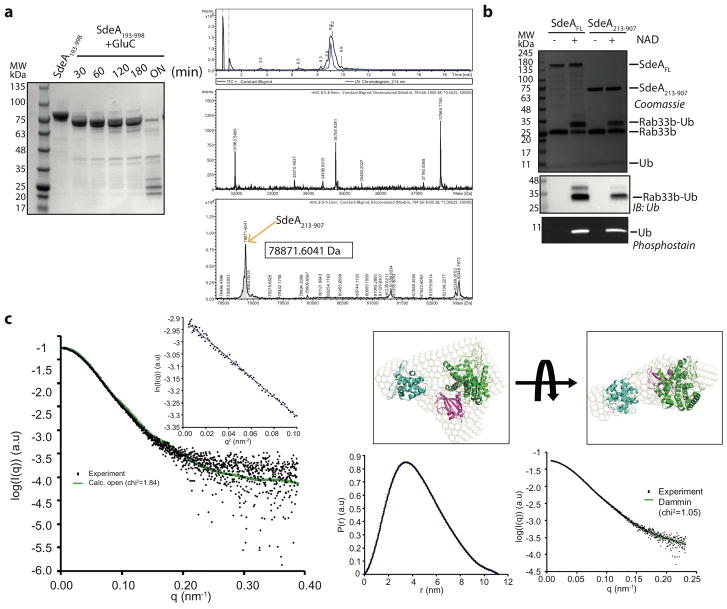
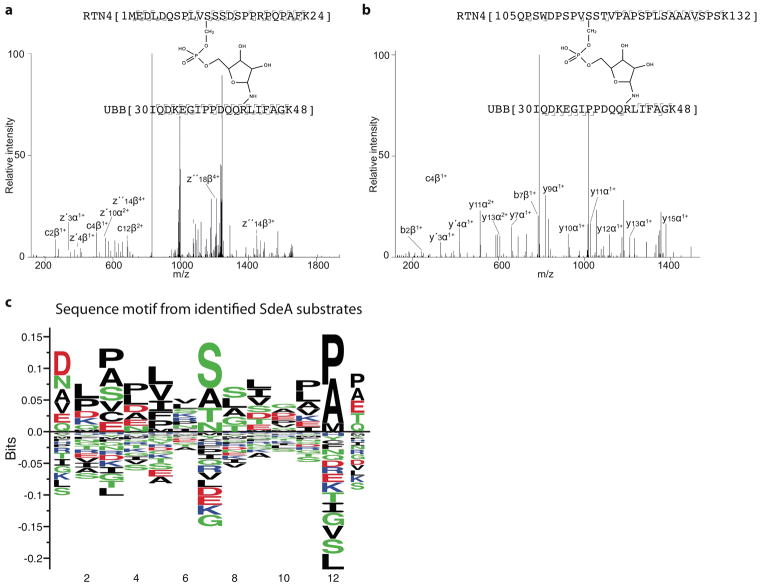
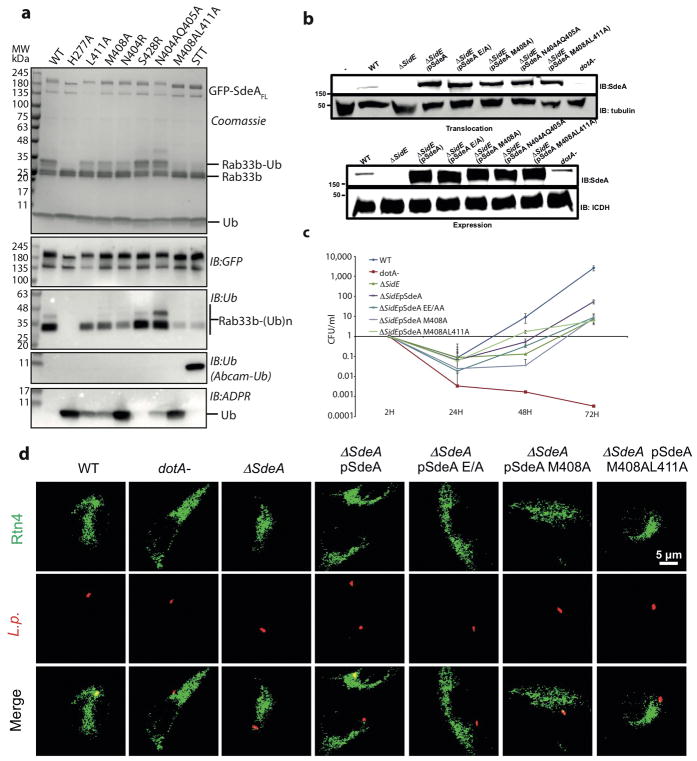
Conventional ubiquitination regulates key cellular processes by catalysing the ATP-dependent formation of an isopeptide bond between ubiquitin (Ub) and primary amines in substrate proteins 1 . Recently, the SidE family of bacterial effector proteins (SdeA, SdeB, SdeC and SidE) from pathogenic Legionella pneumophila were shown to use NAD+ to mediate phosphoribosyl-linked ubiquitination of serine residues in host proteins2, 3. However, the molecular architecture of the catalytic platform that enables this complex multistep process remains unknown. Here we describe the structure of the catalytic core of SdeA, comprising mono-ADP-ribosyltransferase (mART) and phosphodiesterase (PDE) domains, and shed light on the activity of two distinct catalytic sites for serine ubiquitination. The mART catalytic site is composed of an α-helical lobe (AHL) that, together with the mART core, creates a chamber for NAD+ binding and ADP-ribosylation of ubiquitin. The catalytic site in the PDE domain cleaves ADP-ribosylated ubiquitin to phosphoribosyl ubiquitin (PR-Ub) and mediates a two-step PR-Ub transfer reaction: first to a catalytic histidine 277 (forming a transient SdeA H277-PR-Ub intermediate) and subsequently to a serine residue in host proteins. Structural analysis revealed a substrate binding cleft in the PDE domain, juxtaposed with the catalytic site, that is essential for positioning serines for ubiquitination. Using degenerate substrate peptides and newly identified ubiquitination sites in RTN4B, we show that disordered polypeptides with hydrophobic residues surrounding the target serine residues are preferred substrates for SdeA ubiquitination. Infection studies with L. pneumophila expressing substrate-binding mutants of SdeA revealed that substrate ubiquitination, rather than modification of the cellular ubiquitin pool, determines the pathophysiological effect of SdeA during acute bacterial infection.
Conflict of interest statement
The authors declare no competing financial interests.
Figures











Comment in
-
Deciphering the catalytic mechanism of bacterial ubiquitination.Nature. 2018 May;557(7707):644-645. doi: 10.1038/d41586-018-05250-6. Nature. 2018. PMID: 29805175 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
