

Constrained Combinatorial Libraries of Gp2 Proteins Enhance Discovery of PD-L1 Binders
- PMID: 29799714
- PMCID: PMC6051759
- DOI: 10.1021/acscombsci.8b00010
Constrained Combinatorial Libraries of Gp2 Proteins Enhance Discovery of PD-L1 Binders
Abstract
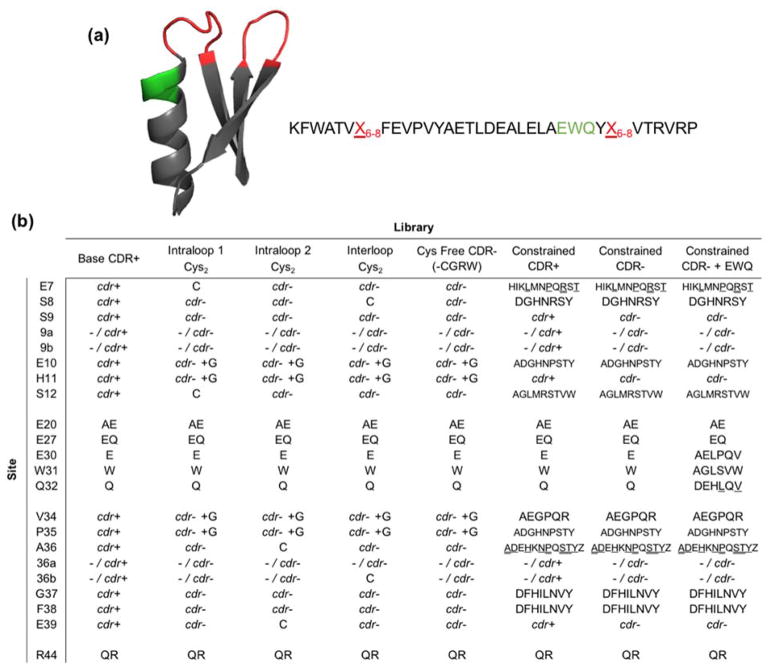
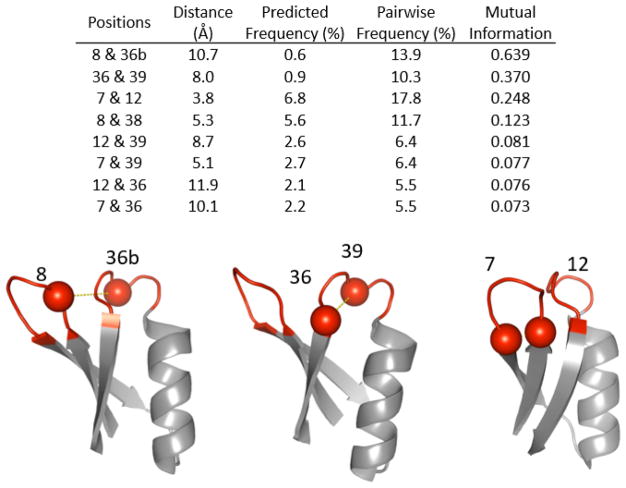
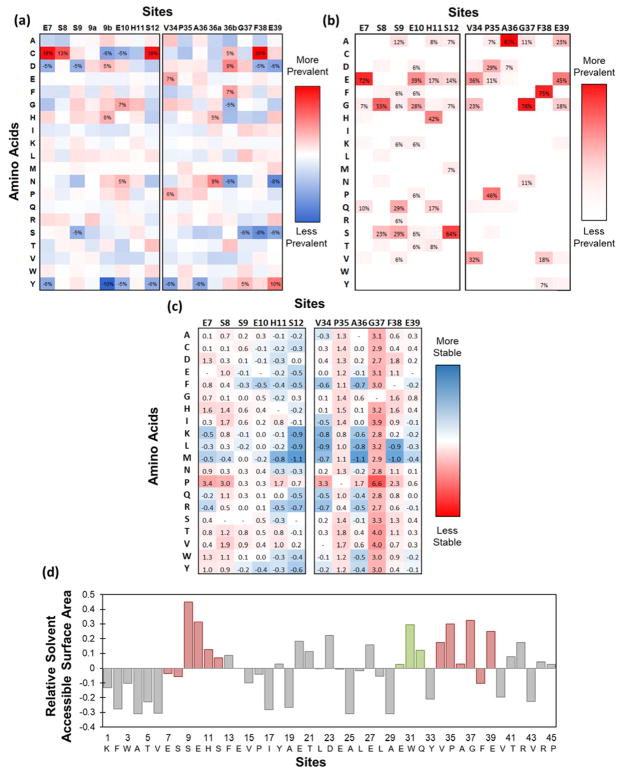
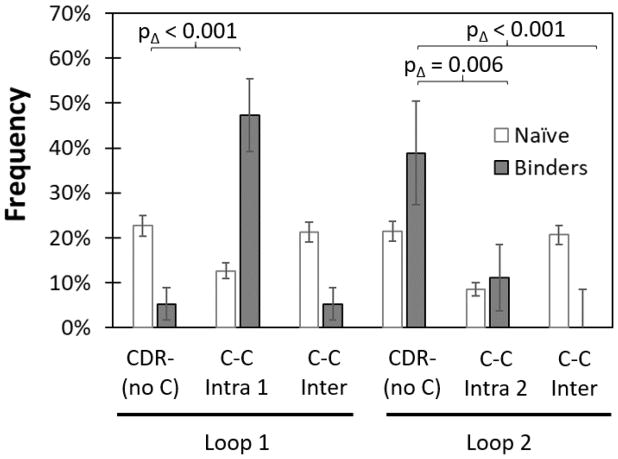
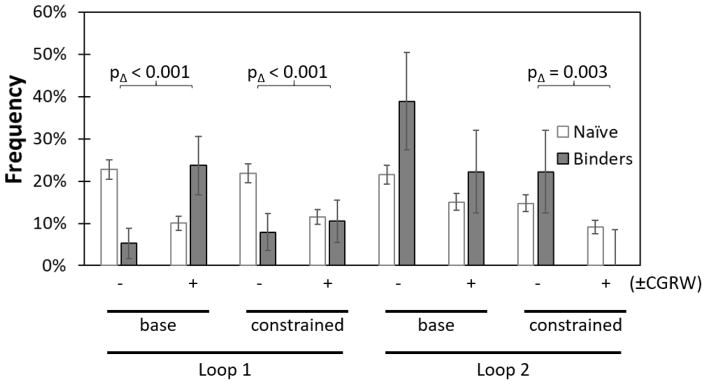
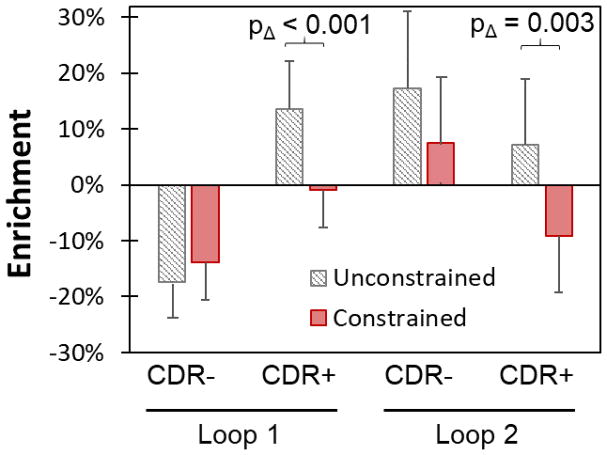
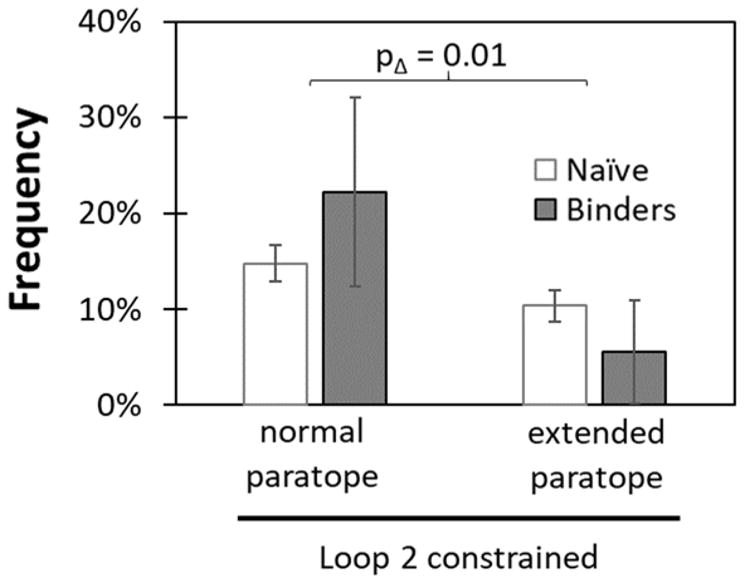
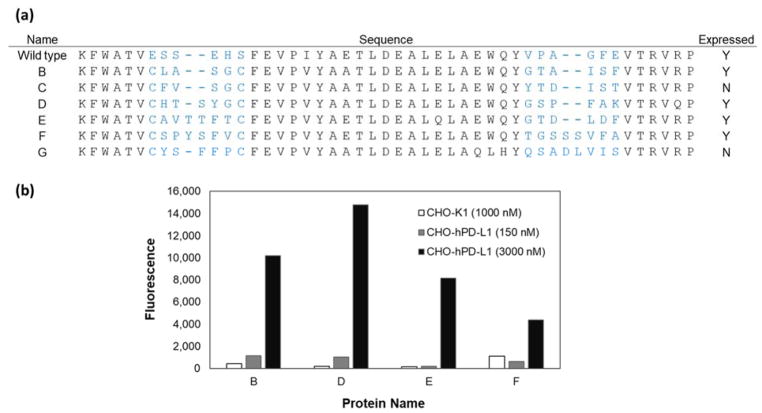
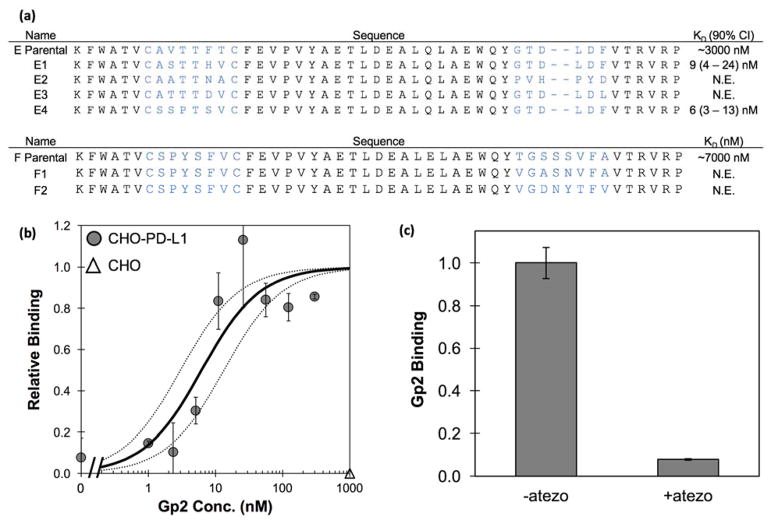
Engineered protein ligands are used for molecular therapy, diagnostics, and industrial biotechnology. The Gp2 domain is a 45-amino acid scaffold that has been evolved for specific, high-affinity binding to multiple targets by diversification of two solvent-exposed loops. Inspired by sitewise enrichment of select amino acids, including cysteine pairs, in earlier Gp2 discovery campaigns, we hypothesized that the breadth and efficiency of de novo Gp2 discovery will be aided by sitewise amino acid constraint within combinatorial library design. We systematically constructed eight libraries and comparatively evaluated their efficacy for binder discovery via yeast display against a panel of targets. Conservation of a cysteine pair at the termini of the first diversified paratope loop increased binder discovery 16-fold ( p < 0.001). Yet two other libraries with conserved cysteine pairs, within the second loop or an interloop pair, did not aid discovery thereby indicating site-specific impact. Via a yeast display protease resistance assay, Gp2 variants from the loop one cysteine pair library were 3.3 ± 2.1-fold ( p = 0.005) more stable than nonconstrained variants. Sitewise constraint of noncysteine residues-guided by previously evolved binders, natural Gp2 homology, computed stability, and structural analysis-did not aid discovery. A panel of binders to programmed death ligand 1 (PD-L1), a key target in cancer immunotherapy, were discovered from the loop 1 cysteine constraint library. Affinity maturation via loop walking resulted in strong, specific cellular PD-L1 affinity ( Kd = 6-9 nM).
Keywords: Gp2 proteins enhance; PD-L1 binders; combinatorial libraries; sitewise amino acid constraint.
Conflict of interest statement
Conflict of Interest: The authors declare no conflict of interest.
Figures

References
-
- Škrlec K, Štrukelj B, Berlec A. Non-immunoglobulin scaffolds: a focus on their targets. Trends Biotechnol. 2015:1–11. - PubMed
-
- Chothia C, Janin J. Principles of protein-protein recognition. Nature. 1975;256:705–708. - PubMed
-
- Kastritis PL, Rodrigues JPGLM, Folkers GE, Boelens R, Bonvin AMJJ. Proteins feel more than they see: fine-tuning of binding affinity by properties of the non-interacting surface. J Mol Biol. 2014;426:2632–52. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
