

A G-protein-coupled chemoattractant receptor recognizes lipopolysaccharide for bacterial phagocytosis
- PMID: 29799847
- PMCID: PMC5969738
- DOI: 10.1371/journal.pbio.2005754
A G-protein-coupled chemoattractant receptor recognizes lipopolysaccharide for bacterial phagocytosis
Abstract
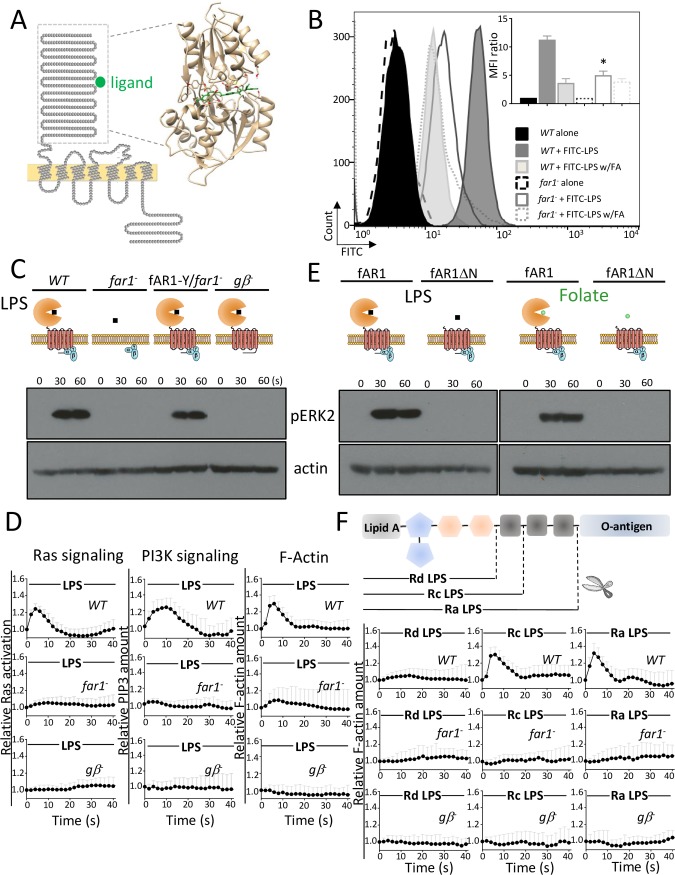
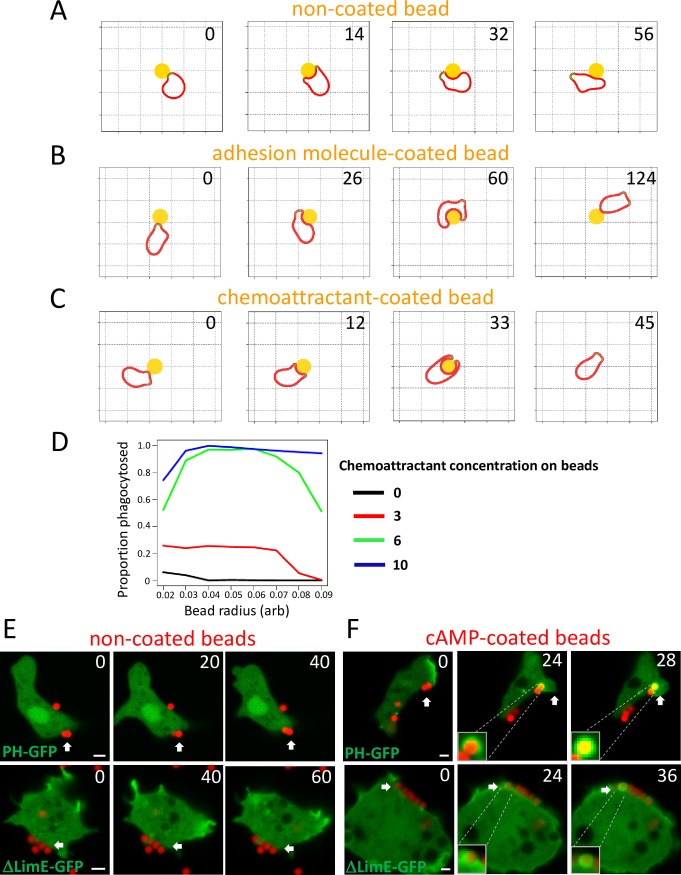
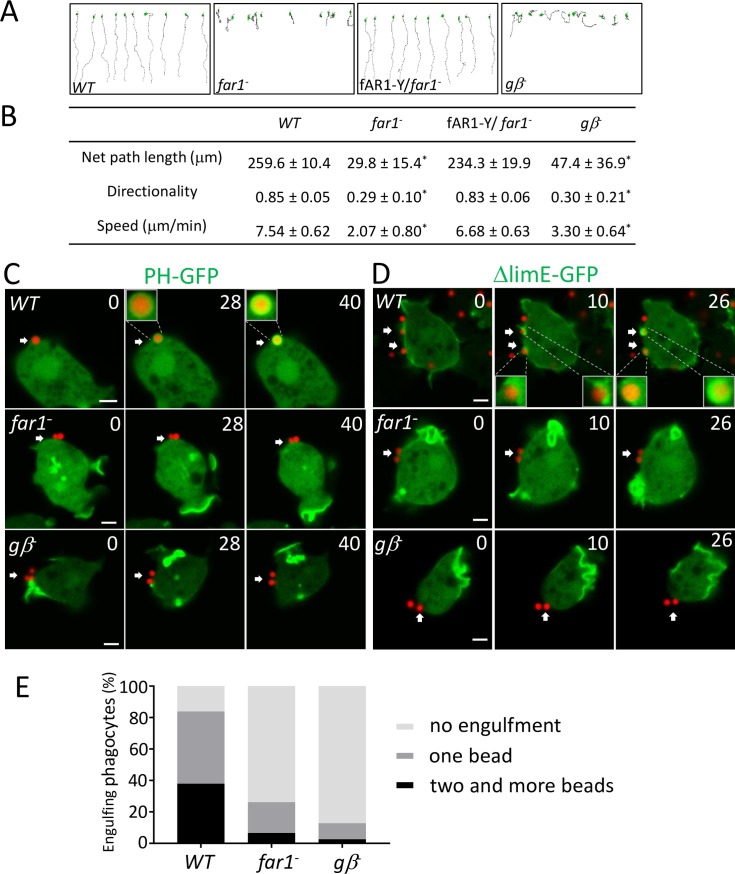
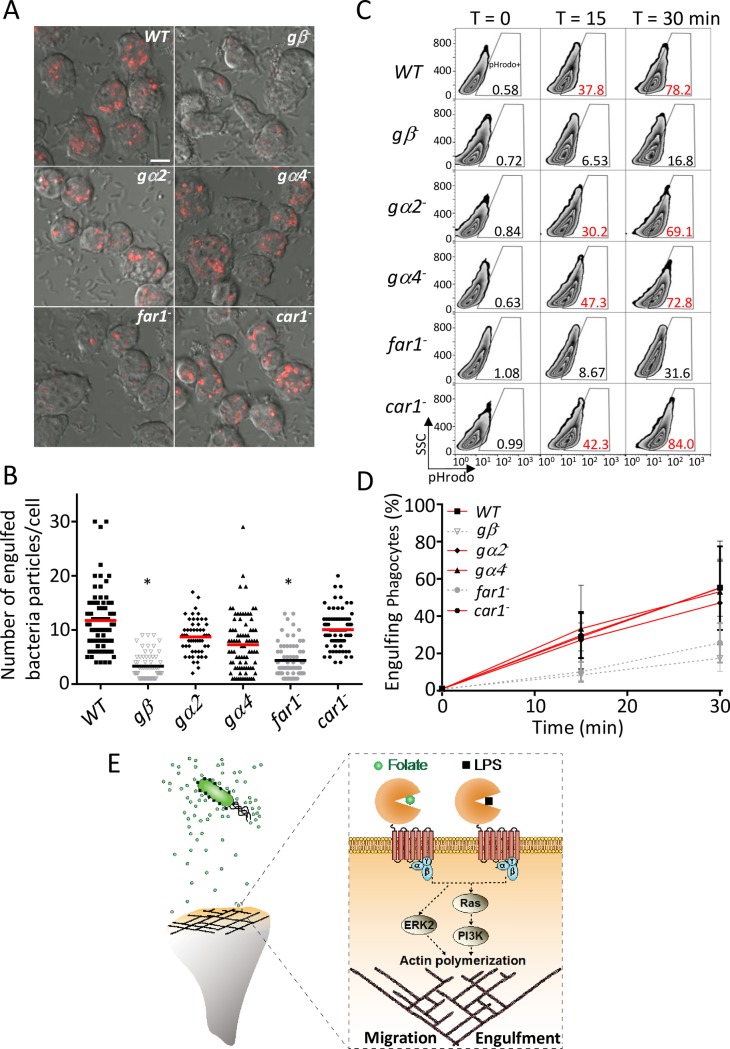
Phagocytes locate microorganisms via chemotaxis and then consume them using phagocytosis. Dictyostelium amoebas are stereotypical phagocytes that prey on diverse bacteria using both processes. However, as typical phagocytic receptors, such as complement receptors or Fcγ receptors, have not been found in Dictyostelium, it remains mysterious how these cells recognize bacteria. Here, we show that a single G-protein-coupled receptor (GPCR), folic acid receptor 1 (fAR1), simultaneously recognizes the chemoattractant folate and the phagocytic cue lipopolysaccharide (LPS), a major component of bacterial surfaces. Cells lacking fAR1 or its cognate G-proteins are defective in chemotaxis toward folate and phagocytosis of Klebsiella aerogenes. Computational simulations combined with experiments show that responses associated with chemotaxis can also promote engulfment of particles coated with chemoattractants. Finally, the extracellular Venus-Flytrap (VFT) domain of fAR1 acts as the binding site for both folate and LPS. Thus, fAR1 represents a new member of the pattern recognition receptors (PRRs) and mediates signaling from both bacterial surfaces and diffusible chemoattractants to reorganize actin for chemotaxis and phagocytosis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Stefater JA 3rd, Ren S, Lang RA, Duffield JS. Metchnikoff's policemen: macrophages in development, homeostasis and regeneration. Trends Mol Med. 2011;17(12):743–52. doi: 10.1016/j.molmed.2011.07.009 ; PubMed Central PMCID: PMC3225647. - DOI - PMC - PubMed
-
- Freeman SA, Grinstein S. Phagocytosis: receptors, signal integration, and the cytoskeleton. Immunol Rev. 2014;262(1):193–215. doi: 10.1111/imr.12212 . - DOI - PubMed
-
- Kaufmann SHE, Dorhoi A. Molecular Determinants in Phagocyte-Bacteria Interactions. Immunity. 2016;44(3):476–91. doi: 10.1016/j.immuni.2016.02.014 . - DOI - PubMed
-
- Rougerie P, Miskolci V, Cox D. Generation of membrane structures during phagocytosis and chemotaxis of macrophages: role and regulation of the actin cytoskeleton. Immunol Rev. 2013;256(1):222–39. doi: 10.1111/imr.12118 ; PubMed Central PMCID: PMC3806206. - DOI - PMC - PubMed
-
- Devreotes PN, Bhattacharya S, Edwards M, Iglesias PA, Lampert T, Miao Y. Excitable Signal Transduction Networks in Directed Cell Migration. Annu Rev Cell Dev Biol. 2017;33:103–25. doi: 10.1146/annurev-cellbio-100616-060739 . - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
