

Conditional Deletion of FOXL2 and SMAD4 in Gonadotropes of Adult Mice Causes Isolated FSH Deficiency
- PMID: 29800110
- PMCID: PMC6692885
- DOI: 10.1210/en.2018-00100
Conditional Deletion of FOXL2 and SMAD4 in Gonadotropes of Adult Mice Causes Isolated FSH Deficiency
Abstract
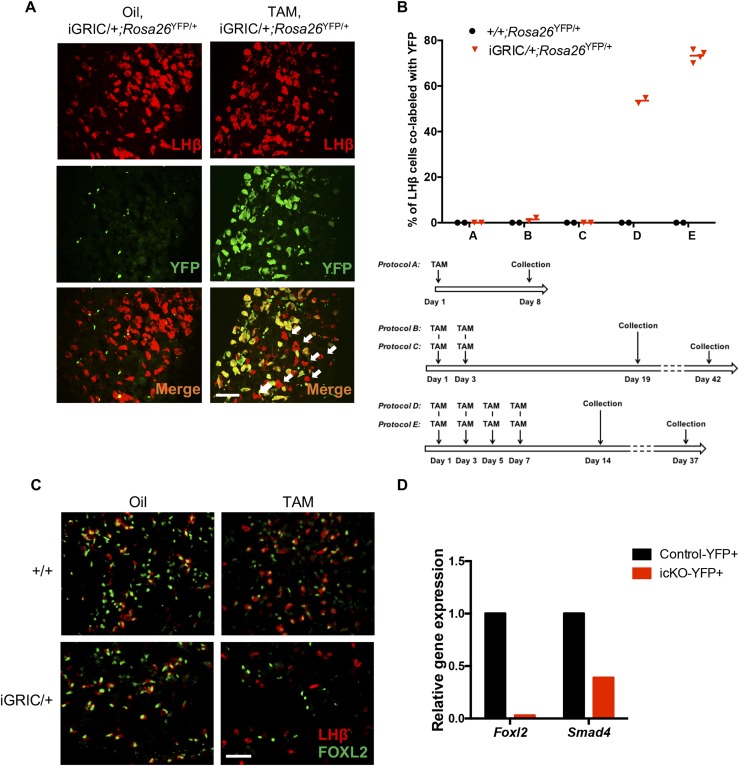
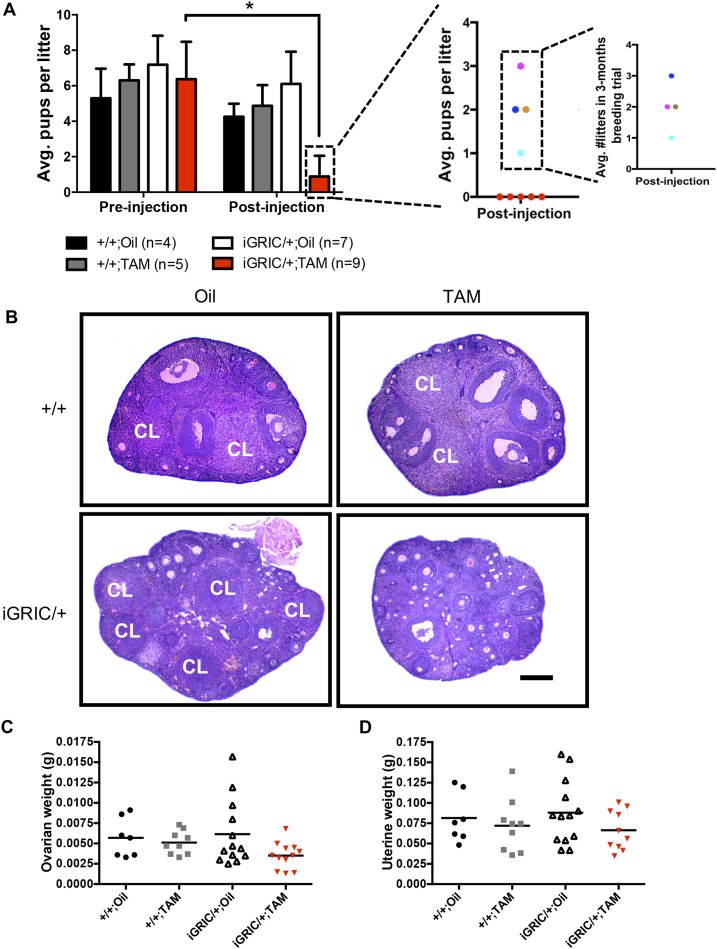
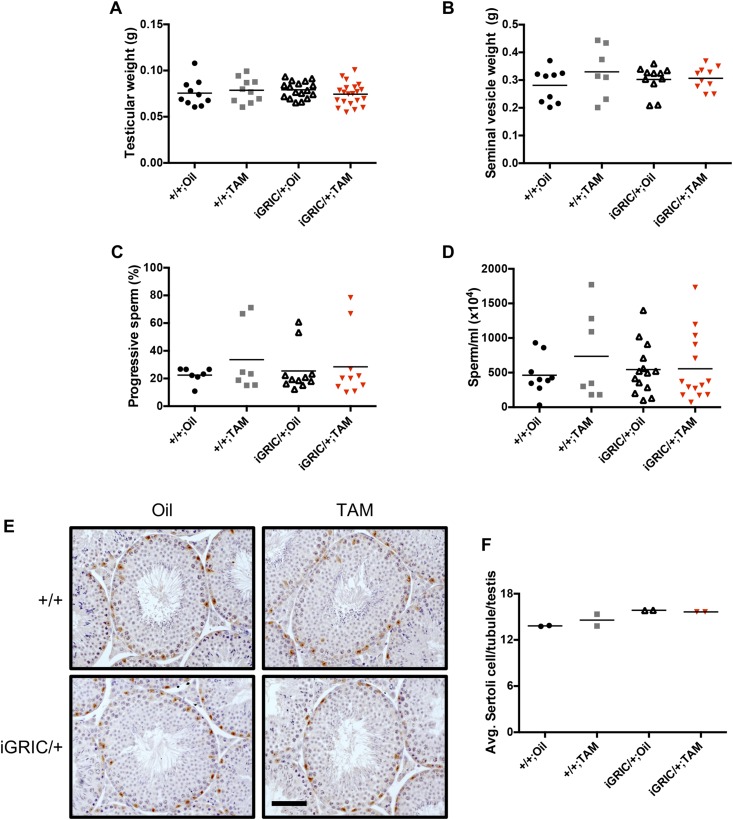
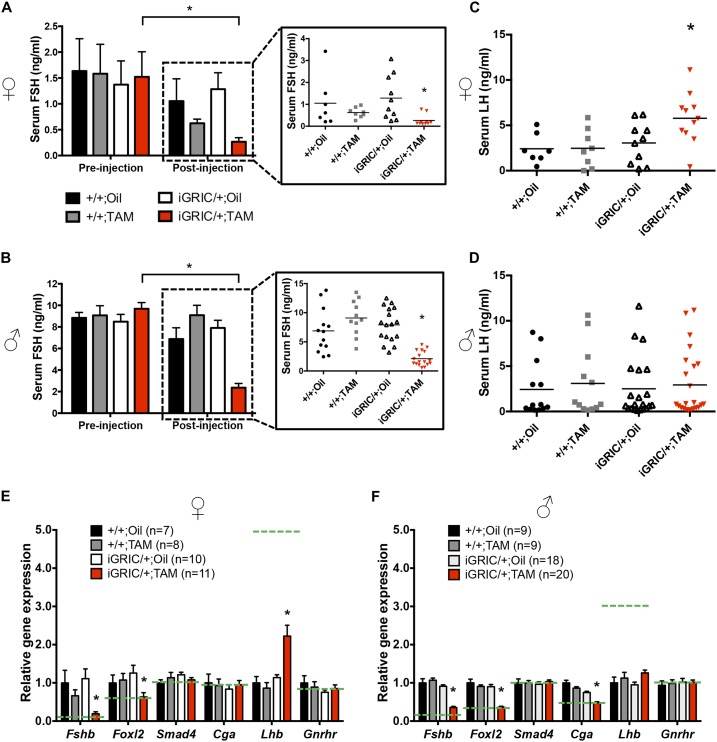
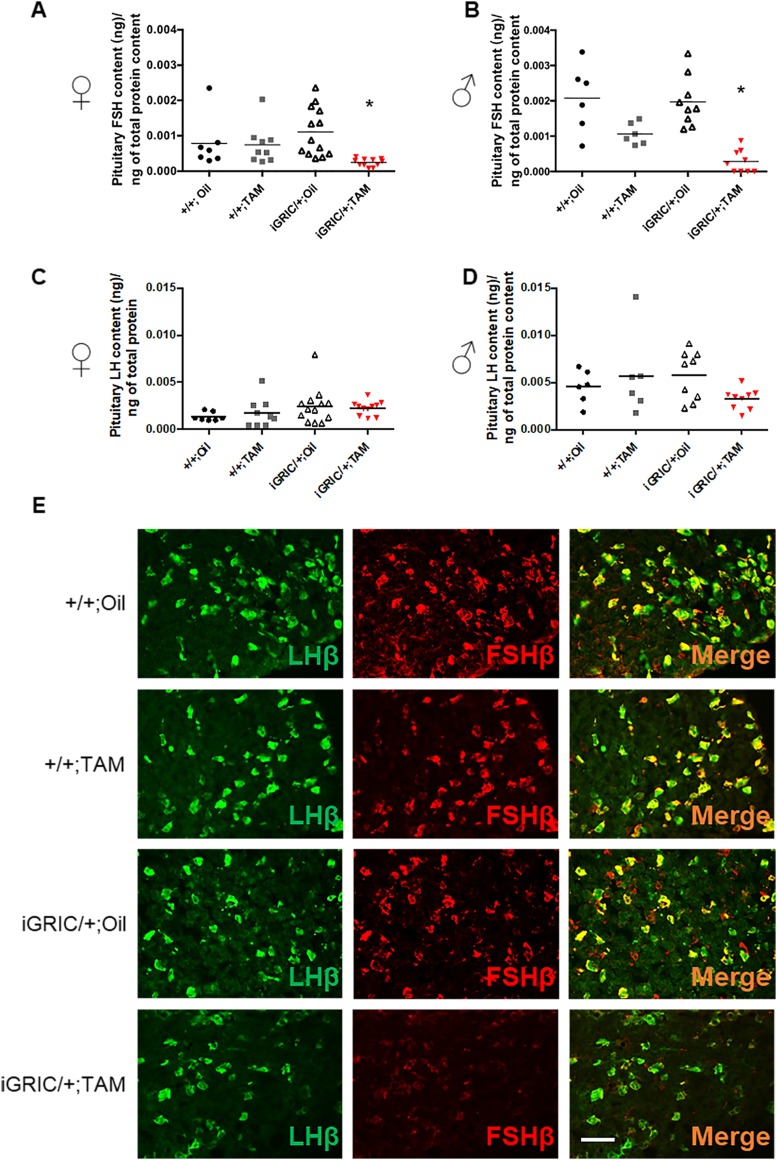
The glycoprotein FSH, a product of pituitary gonadotrope cells, regulates ovarian follicle development in females and spermatogenesis in males. FSH is a heterodimer of the common α gonadotropin subunit and the hormone-specific FSHβ subunit (a product of the Fshb gene). Using a conditional knockout approach (Cre-lox), we previously demonstrated that Fshb expression in mice depends on the transcription factors forkhead box L2 (FOXL2) and SMAD4. Deletion of Foxl2 or Smad4 alone led to FSH deficiency, female subfertility, and oligozoospermia in males. Simultaneous deletion of the two genes yielded a greater suppression of FSH and female sterility. The Cre-driver used previously was first active during embryonic development. Therefore, it is unclear whether FOXL2 and SMAD4 play important roles in the development or adult function of gonadotropes, or both. To address this question, we developed a tamoxifen-inducible Cre-driver line, which enabled Foxl2 and Smad4 gene deletions in gonadotropes of adult mice. After tamoxifen treatment, females with previously demonstrated fertility exhibited profound reductions in FSH levels, arrested ovarian follicle development, and sterility. FSH levels were comparably reduced in males 1 or 2 months after treatment; however, spermatogenesis was unaffected. These data indicate that (1) FOXL2 and SMAD4 are necessary to maintain FSH synthesis in gonadotrope cells of adult mice, (2) FSH is essential for female reproduction but appears to be unnecessary for the maintenance of spermatogenesis in adult male mice, and (3) the inducible Cre-driver line developed here provides a powerful tool to interrogate gene function in gonadotrope cells of adult mice.
Figures

Similar articles
-
Human Follicle-Stimulating Hormone ß Subunit Expression Depends on FOXL2 and SMAD4.Endocrinology. 2020 May 1;161(5):bqaa045. doi: 10.1210/endocr/bqaa045. Endocrinology. 2020. PMID: 32191302 Free PMC article.
-
SMAD3 Regulates Follicle-stimulating Hormone Synthesis by Pituitary Gonadotrope Cells in Vivo.J Biol Chem. 2017 Feb 10;292(6):2301-2314. doi: 10.1074/jbc.M116.759167. Epub 2016 Dec 19. J Biol Chem. 2017. PMID: 27994055 Free PMC article.
-
Impaired fertility and FSH synthesis in gonadotrope-specific Foxl2 knockout mice.Mol Endocrinol. 2013 Mar;27(3):407-21. doi: 10.1210/me.2012-1286. Epub 2013 Jan 22. Mol Endocrinol. 2013. PMID: 23340250 Free PMC article.
-
Minireview: Activin Signaling in Gonadotropes: What Does the FOX say… to the SMAD?Mol Endocrinol. 2015 Jul;29(7):963-77. doi: 10.1210/me.2015-1004. Epub 2015 May 5. Mol Endocrinol. 2015. PMID: 25942106 Free PMC article. Review.
-
Fox tales: regulation of gonadotropin gene expression by forkhead transcription factors.Mol Cell Endocrinol. 2014 Mar 25;385(1-2):62-70. doi: 10.1016/j.mce.2013.09.034. Epub 2013 Oct 4. Mol Cell Endocrinol. 2014. PMID: 24099863 Free PMC article. Review.
Cited by
-
Smad4 and FoxH1 potentially interact to regulate cyp19a1a promoter in the ovary of ricefield eel (Monopterus albus).Biol Sex Differ. 2024 Jul 30;15(1):60. doi: 10.1186/s13293-024-00636-w. Biol Sex Differ. 2024. PMID: 39080808 Free PMC article.
-
Rationale and Roadmap for Developing Panels of Hotspot Cancer Driver Gene Mutations as Biomarkers of Cancer Risk.Environ Mol Mutagen. 2020 Jan;61(1):152-175. doi: 10.1002/em.22326. Epub 2019 Oct 6. Environ Mol Mutagen. 2020. PMID: 31469467 Free PMC article. Review.
-
Gonadotrope-specific deletion of the BMP type 2 receptor does not affect reproductive physiology in mice†‡.Biol Reprod. 2020 Mar 13;102(3):639-646. doi: 10.1093/biolre/ioz206. Biol Reprod. 2020. PMID: 31724029 Free PMC article.
-
Addition of an N-Terminal Poly-Glutamate Fusion Tag Improves Solubility and Production of Recombinant TAT-Cre Recombinase in Escherichia coli.J Microbiol Biotechnol. 2020 Jan 28;30(1):109-117. doi: 10.4014/jmb.1909.09028. J Microbiol Biotechnol. 2020. PMID: 31693834 Free PMC article.
-
Selected Genetic Factors Associated with Primary Ovarian Insufficiency.Int J Mol Sci. 2023 Feb 23;24(5):4423. doi: 10.3390/ijms24054423. Int J Mol Sci. 2023. PMID: 36901862 Free PMC article. Review.
References
-
- Kumar TR, Wang Y, Lu N, Matzuk MM. Follicle stimulating hormone is required for ovarian follicle maturation but not male fertility. Nat Genet. 1997;15(2):201–204. - PubMed
-
- Matthews CH, Borgato S, Beck-Peccoz P, Adams M, Tone Y, Gambino G, Casagrande S, Tedeschini G, Benedetti A, Chatterjee VK. Primary amenorrhoea and infertility due to a mutation in the beta-subunit of follicle-stimulating hormone. Nat Genet. 1993;5(1):83–86. - PubMed
-
- Richards JS. Hormonal control of gene expression in the ovary. Endocr Rev. 1994;15(6):725–751. - PubMed
-
- Couzinet B, Lestrat N, Brailly S, Forest M, Schaison G. Stimulation of ovarian follicular maturation with pure follicle-stimulating hormone in women with gonadotropin deficiency. J Clin Endocrinol Metab. 1988;66(3):552–556. - PubMed
-
- Singh J, Handelsman DJ. Neonatal administration of FSH increases Sertoli cell numbers and spermatogenesis in gonadotropin-deficient (hpg) mice. J Endocrinol. 1996;151(1):37–48. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
