

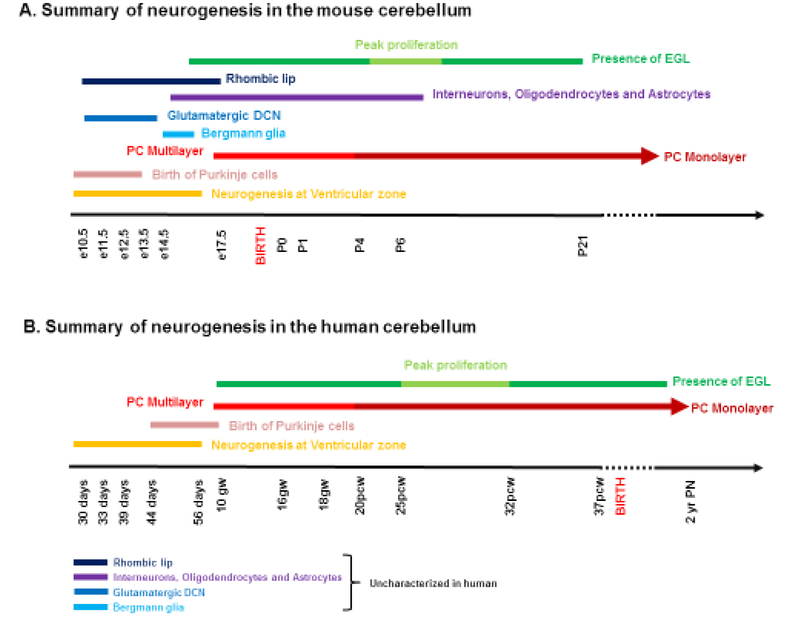
What cerebellar malformations tell us about cerebellar development
- PMID: 29802918
- PMCID: PMC6240394
- DOI: 10.1016/j.neulet.2018.05.032
What cerebellar malformations tell us about cerebellar development
Abstract
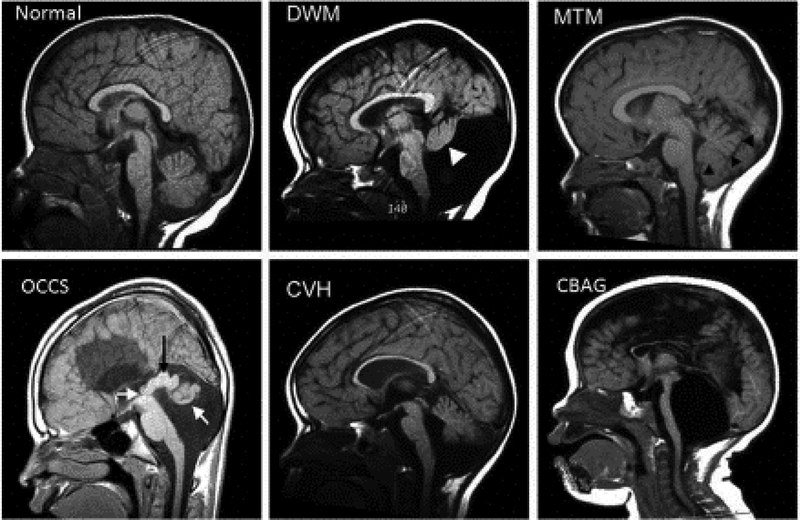
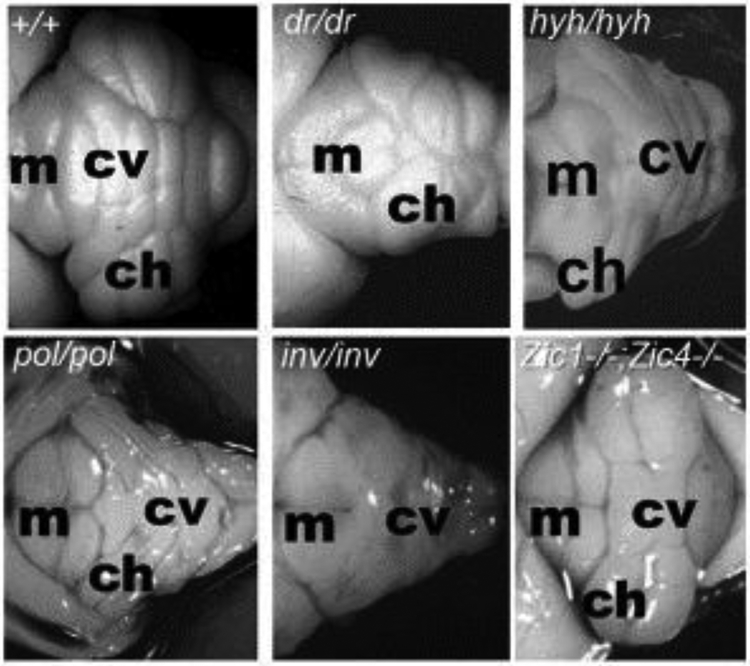
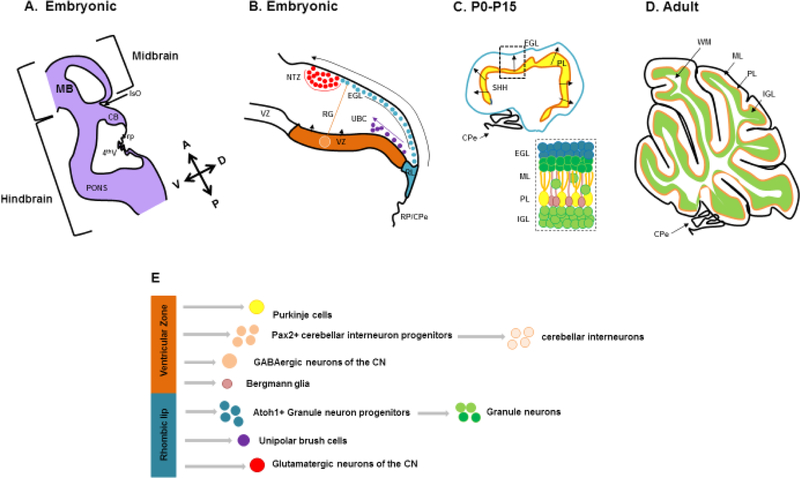
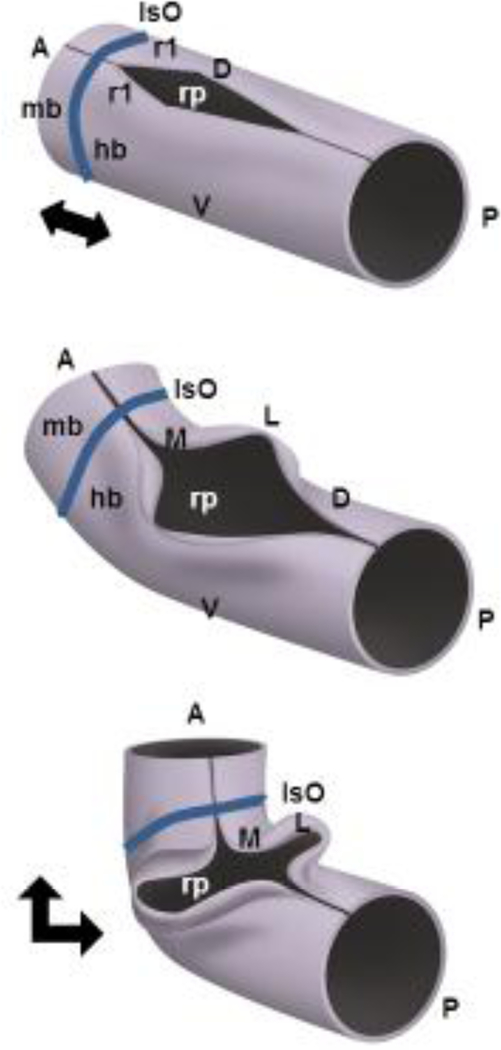
Structural birth defects of the cerebellum, or cerebellar malformations, in humans, have long been recognized. However, until recently there has been little progress in elucidating their developmental pathogenesis. Innovations in brain imaging and human genetic technologies over the last 2 decades have led to better classifications of these disorders and identification of several causative genes. In contrast, cerebellar malformations in model organisms, particularly mice, have been the focus of intense study for more than 70 years. As a result, many of the molecular, genetic and cellular programs that drive formation of the cerebellum have been delineated in mice. In this review, we overview the basic epochs and key molecular regulators of the developmental programs that build the structure of the mouse cerebellum. This mouse-centric approach has been a useful to interpret the developmental pathogenesis of human cerebellar malformations. However, it is becoming apparent that we actually know very little regarding the specifics of human cerebellar development beyond what is inferred from mice. A better understanding of human cerebellar development will not only facilitate improved diagnosis of human cerebellar malformations, but also lead to the development of treatment paradigms for these important neurodevelopmental disorders.
Keywords: Cerebellar malformation; Development; Human; Model organism; Mouse; Neurogenetics; Pathogenesis.
Copyright © 2018 Elsevier B.V. All rights reserved.
Figures
References
-
- Baptista CA, Hatten ME, Blazeski R and Mason CA (1994) ‘Cell-cell interactions influence survival and differentiation of purified Purkinje cells in vitro’, Neuron 12(2): 243–60. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
