

Synaptic activity induces input-specific rearrangements in a targeted synaptic protein interaction network
- PMID: 29804286
- PMCID: PMC6150823
- DOI: 10.1111/jnc.14466
Synaptic activity induces input-specific rearrangements in a targeted synaptic protein interaction network
Abstract
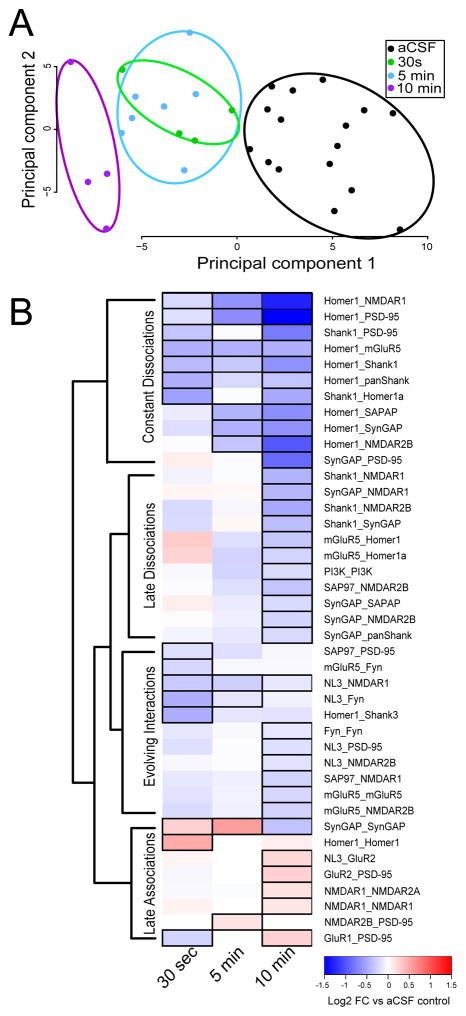
Cells utilize dynamic, network-level rearrangements in highly interconnected protein interaction networks to transmit and integrate information from distinct signaling inputs. Despite the importance of protein interaction network dynamics, the organizational logic underlying information flow through these networks is not well understood. Previously, we developed the quantitative multiplex co-immunoprecipitation platform, which allows for the simultaneous and quantitative measurement of the amount of co-association between large numbers of proteins in shared complexes. Here, we adapt quantitative multiplex co-immunoprecipitation to define the activity-dependent dynamics of an 18-member protein interaction network in order to better understand the underlying principles governing glutamatergic signal transduction. We first establish that immunoprecipitation detected by flow cytometry can detect activity-dependent changes in two known protein-protein interactions (Homer1-mGluR5 and PSD-95-SynGAP). We next demonstrate that neuronal stimulation elicits a coordinated change in our targeted protein interaction network, characterized by the initial dissociation of Homer1 and SynGAP-containing complexes followed by increased associations among glutamate receptors and PSD-95. Finally, we show that stimulation of distinct glutamate receptor types results in different modular sets of protein interaction network rearrangements, and that cells activate both modules in order to integrate complex inputs. This analysis demonstrates that cells respond to distinct types of glutamatergic input by modulating different combinations of protein co-associations among a targeted network of proteins. Our data support a model of synaptic plasticity in which synaptic stimulation elicits dissociation of pre-existing multiprotein complexes, opening binding slots in scaffold proteins and allowing for the recruitment of additional glutamatergic receptors. Open Science: This manuscript was awarded with the Open Materials Badge. For more information see: https://cos.io/our-services/open-science-badges/.
Keywords: SynGAP; glutamate signaling; homer; protein interaction network; quantitative multiplex immunoprecipitation; synapse.
© 2018 International Society for Neurochemistry.
Figures






References
Works Cited
-
- Bashir ZI, Bortolotto ZA, Davies CH, et al. Induction of LTP in the hippocampus needs synaptic activation of glutamate metabotropic receptors. Nature. 1993;363:347–350. - PubMed
-
- Beattie EC, Carroll RC, Yu X, Morishita W, Yasuda H, von Zastrow M, Malenka RC. Regulation of AMPA receptor endocytosis by a signaling mechanism shared with LTD. Nat Neurosci. 2000;3:1291–1300. - PubMed
-
- Cao FF, Xu LM, Zhang X, Wang Y, Li MQ, Uzan G, Peng B, Zhang DH. IP-FCM platform detects the existence and regulator-caused dissociation of components in naturally assembled HSP90 complex. Cytometry A. 2014;85:359–367. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
