

Structural Basis of Smoothened Activation in Hedgehog Signaling
- PMID: 29804838
- PMCID: PMC6046275
- DOI: 10.1016/j.cell.2018.04.029
Structural Basis of Smoothened Activation in Hedgehog Signaling
Erratum in
-
Structural Basis of Smoothened Activation in Hedgehog Signaling.Cell. 2018 Sep 20;175(1):295-297. doi: 10.1016/j.cell.2018.09.003. Cell. 2018. PMID: 30241610 Free PMC article. No abstract available.
Abstract
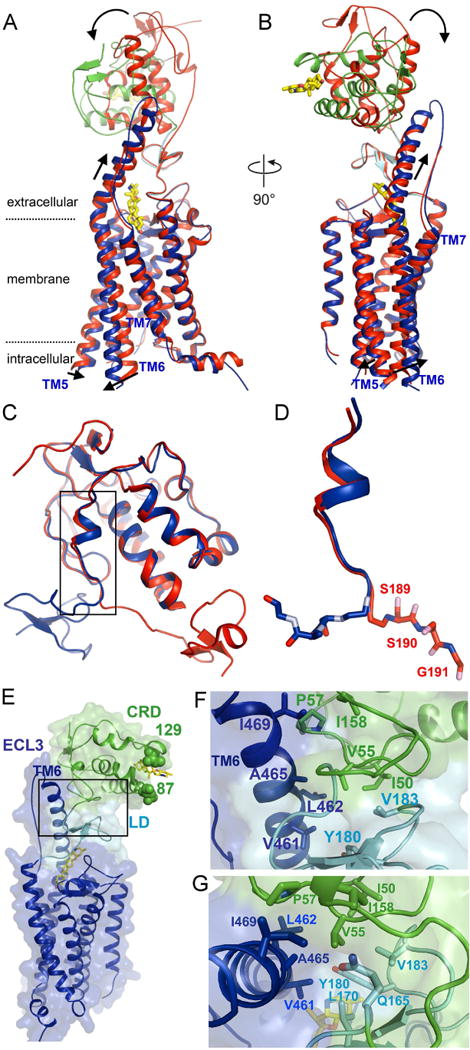
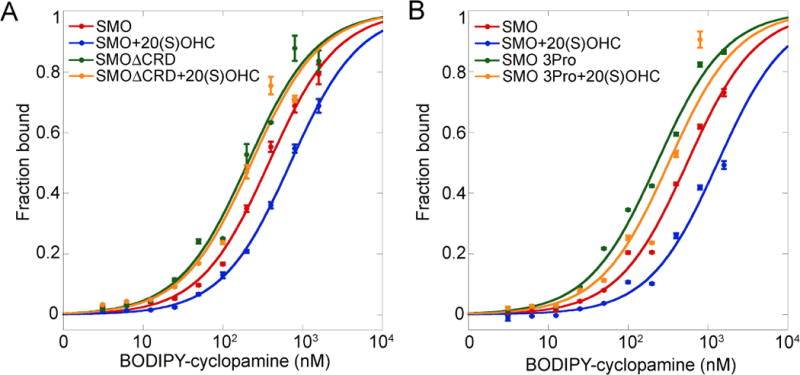
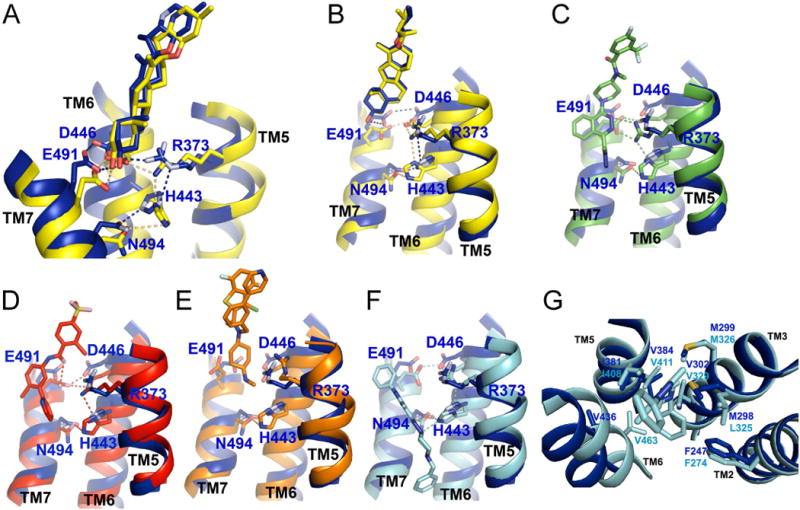
The seven-transmembrane-spanning protein Smoothened is the central transducer in Hedgehog signaling, a pathway fundamental in development and in cancer. Smoothened is activated by cholesterol binding to its extracellular cysteine-rich domain (CRD). How this interaction leads to changes in the transmembrane domain and Smoothened activation is unknown. Here, we report crystal structures of sterol-activated Smoothened. The CRD undergoes a dramatic reorientation, allosterically causing the transmembrane domain to adopt a conformation similar to active G-protein-coupled receptors. We show that Smoothened contains a unique inhibitory π-cation lock, which is broken on activation and is disrupted in constitutively active oncogenic mutants. Smoothened activation opens a hydrophobic tunnel, suggesting a pathway for cholesterol movement from the inner membrane leaflet to the CRD. All Smoothened antagonists bind the transmembrane domain and block tunnel opening, but cyclopamine also binds the CRD, inducing the active transmembrane conformation. Together, these results define the mechanisms of Smoothened activation and inhibition.
Keywords: Hedgehog; Smoothened; cancer; cholesterol; cilia; membrane; oncogene; signaling.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Alcedo J, Ayzenzon M, Von Ohlen T, Noll M, Hooper JE. The Drosophila smoothened gene encodes a seven-pass membrane protein, a putative receptor for the hedgehog signal. Cell. 1996;86:221–232. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
