

Mutations in the Epithelial Cadherin-p120-Catenin Complex Cause Mendelian Non-Syndromic Cleft Lip with or without Cleft Palate
- PMID: 29805042
- PMCID: PMC5992119
- DOI: 10.1016/j.ajhg.2018.04.009
Mutations in the Epithelial Cadherin-p120-Catenin Complex Cause Mendelian Non-Syndromic Cleft Lip with or without Cleft Palate
Abstract
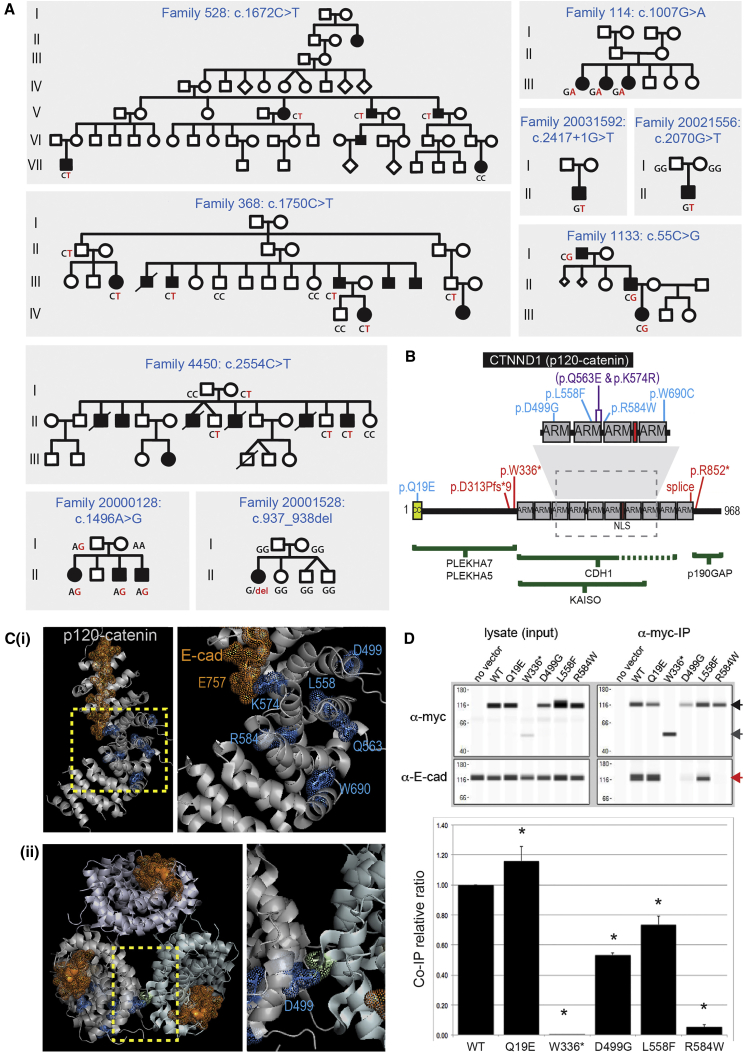
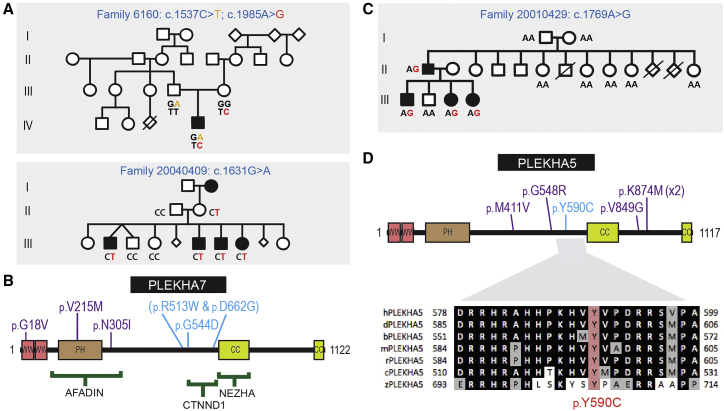
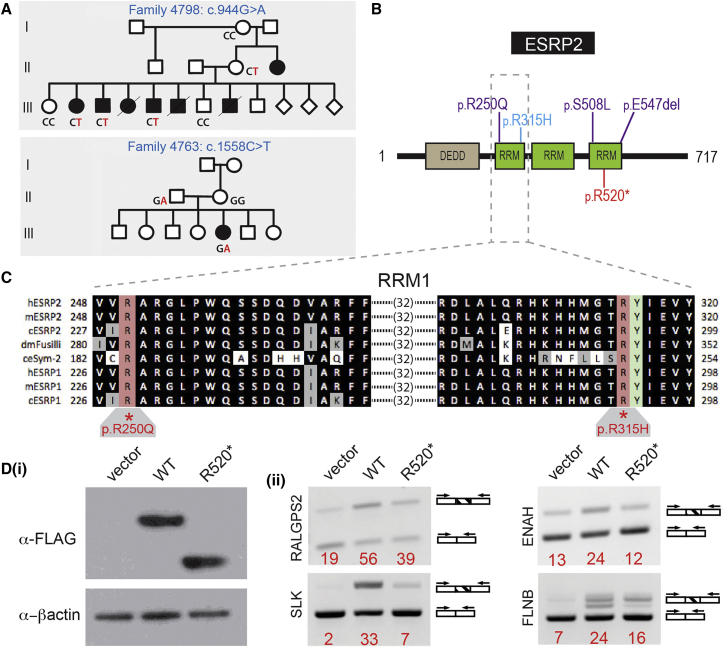
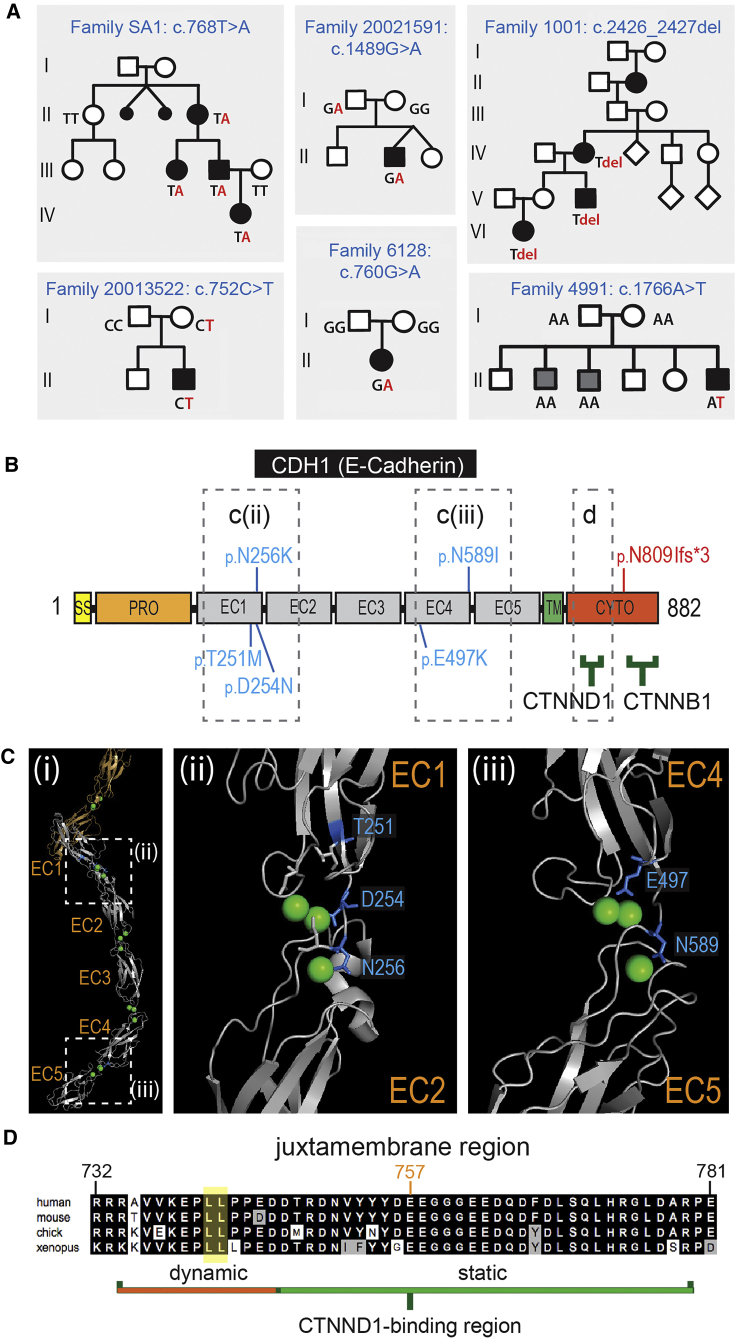
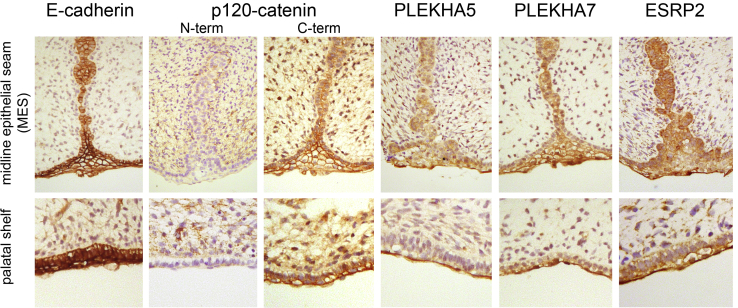
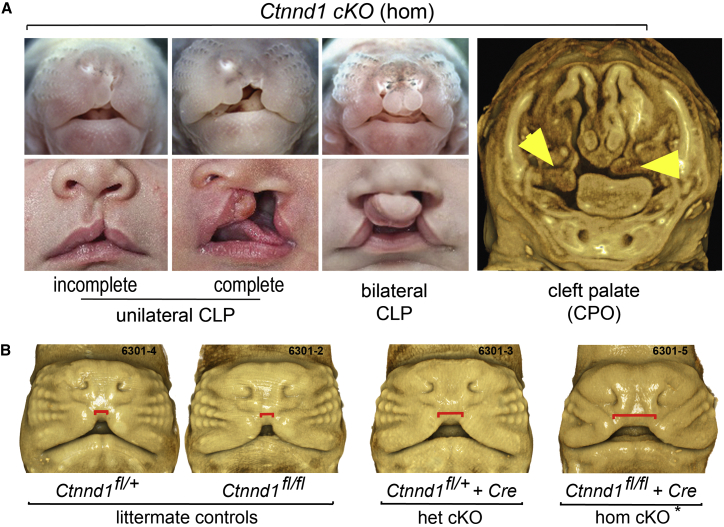
Non-syndromic cleft lip with or without cleft palate (NS-CL/P) is one of the most common human birth defects and is generally considered a complex trait. Despite numerous loci identified by genome-wide association studies, the effect sizes of common variants are relatively small, with much of the presumed genetic contribution remaining elusive. We report exome-sequencing results in 209 people from 72 multi-affected families with pedigree structures consistent with autosomal-dominant inheritance and variable penetrance. Herein, pathogenic variants are described in four genes encoding components of the p120-catenin complex (CTNND1, PLEKHA7, PLEKHA5) and an epithelial splicing regulator (ESRP2), in addition to the known CL/P-associated gene, CDH1, which encodes E-cadherin. The findings were also validated in a second cohort of 497 people with NS-CL/P, comprising small families and singletons with pathogenic variants in these genes identified in 14% of multi-affected families and 2% of the replication cohort of smaller families. Enriched expression of each gene/protein in human and mouse embryonic oro-palatal epithelia, demonstration of functional impact of CTNND1 and ESRP2 variants, and recapitulation of the CL/P spectrum in Ctnnd1 knockout mice support a causative role in CL/P pathogenesis. These data show that primary defects in regulators of epithelial cell adhesion are the most significant contributors to NS-CL/P identified to date and that inherited and de novo single gene variants explain a substantial proportion of NS-CL/P.
Keywords: adherens junction; cadherin; catenin; cell adhesion; cleft lip; cleft lip/palate; cleft palate; epithelia; exome sequencing; knockout.
Copyright © 2018 American Society of Human Genetics. All rights reserved.
Figures
References
-
- Ghoumid J., Stichelbout M., Jourdain A.S., Frenois F., Lejeune-Dumoulin S., Alex-Cordier M.P., Lebrun M., Guerreschi P., Duquennoy-Martinot V., Vinchon M. Blepharocheilodontic syndrome is a CDH1 pathway-related disorder due to mutations in CDH1 and CTNND1. Genet. Med. 2017;19:1013–1021. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
