

A Role for Hypocretin/Orexin in Metabolic and Sleep Abnormalities in a Mouse Model of Non-metastatic Breast Cancer
- PMID: 29805100
- PMCID: PMC6031468
- DOI: 10.1016/j.cmet.2018.04.021
A Role for Hypocretin/Orexin in Metabolic and Sleep Abnormalities in a Mouse Model of Non-metastatic Breast Cancer
Abstract
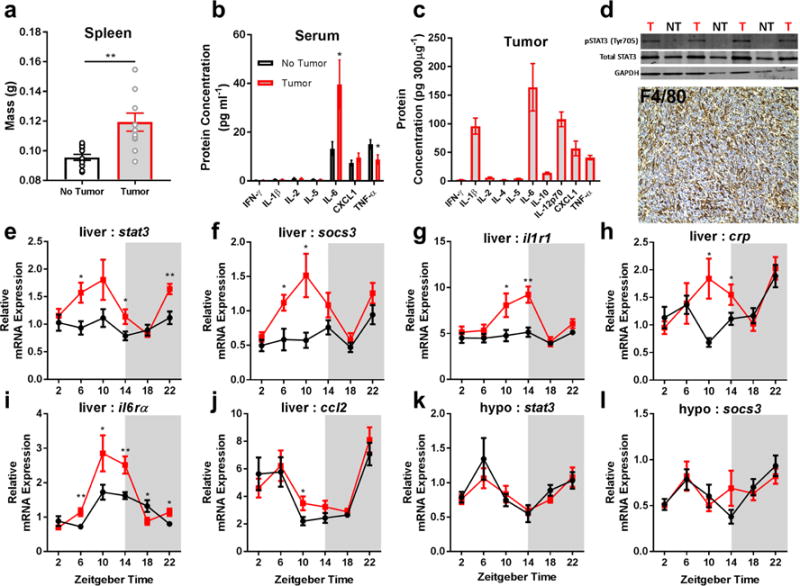
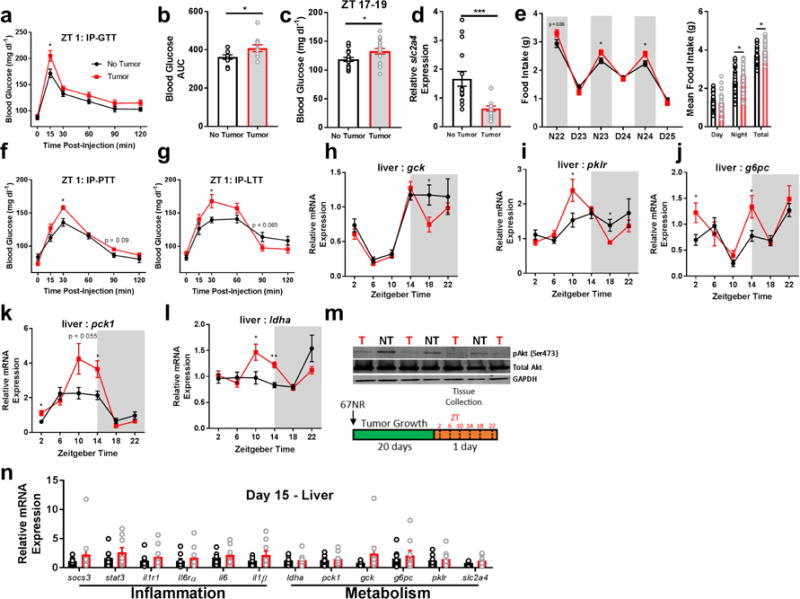
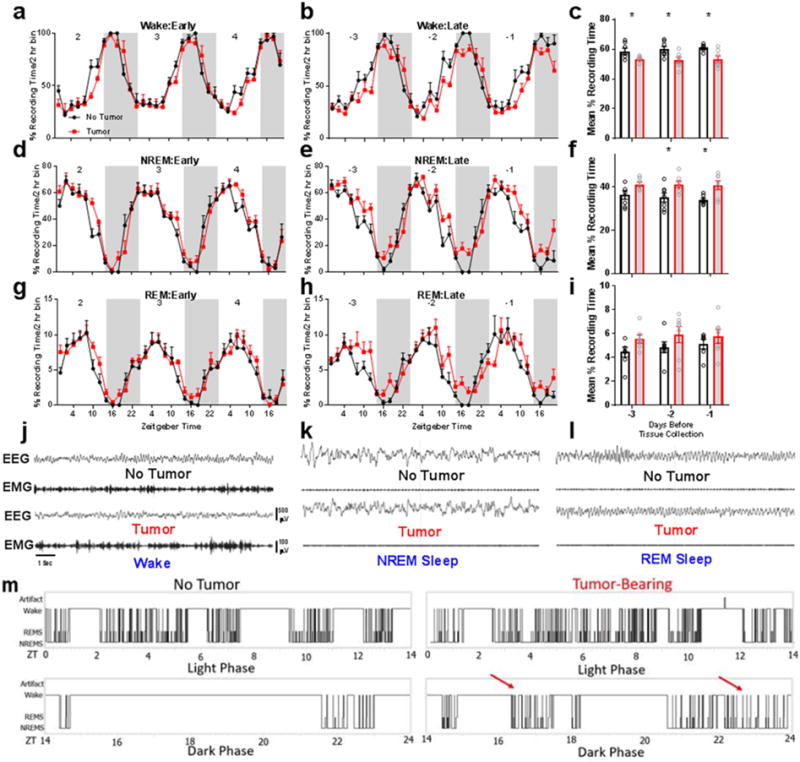
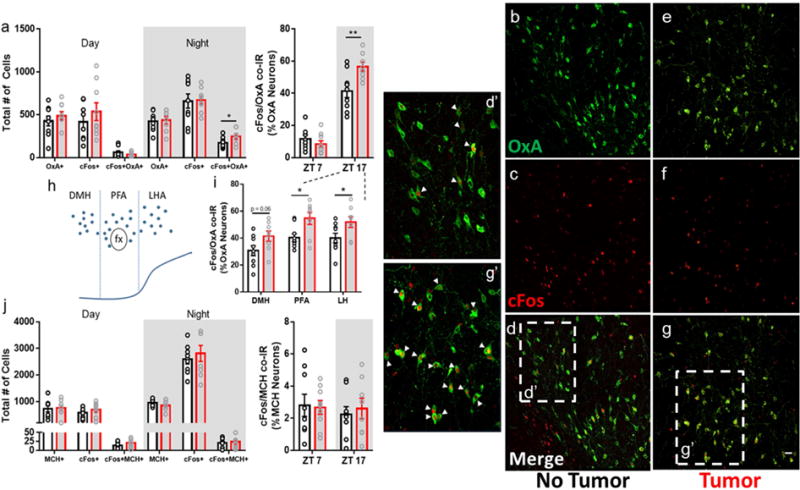
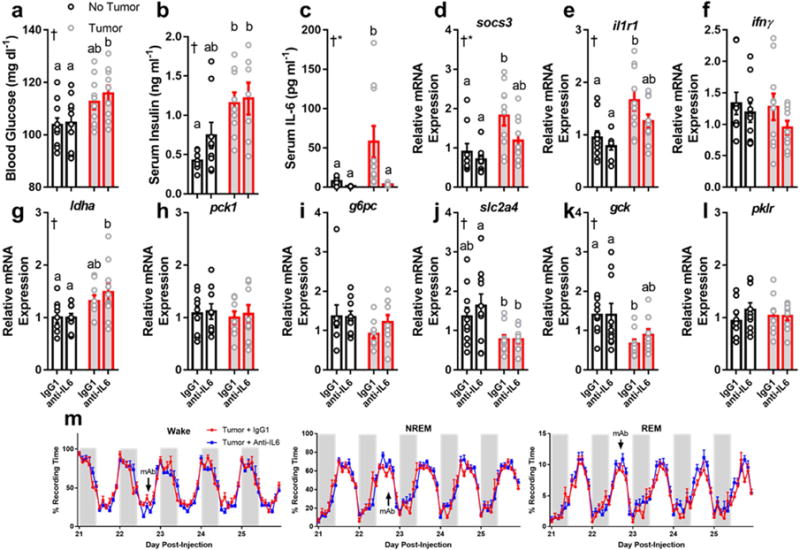
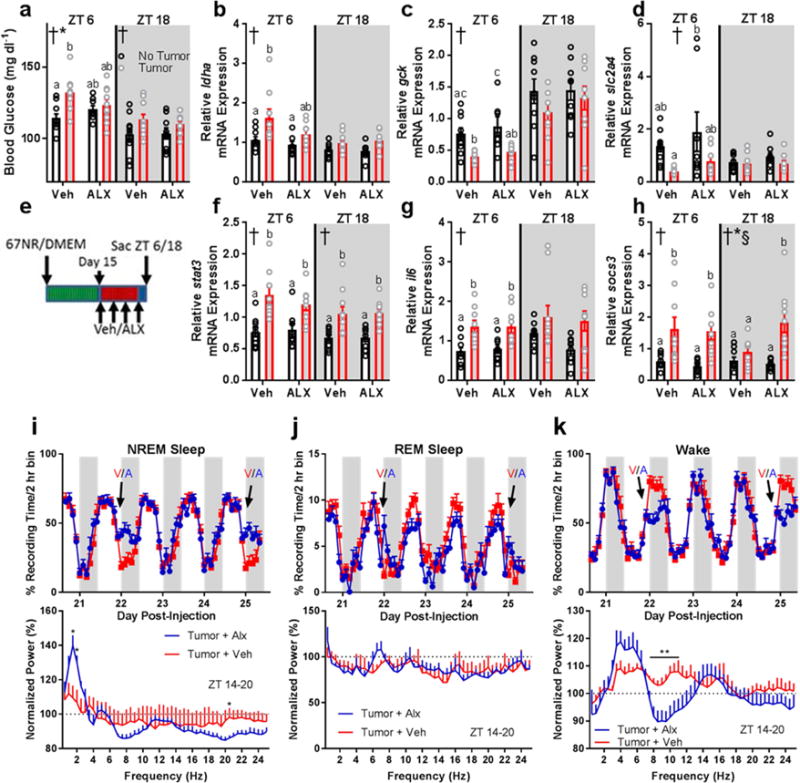
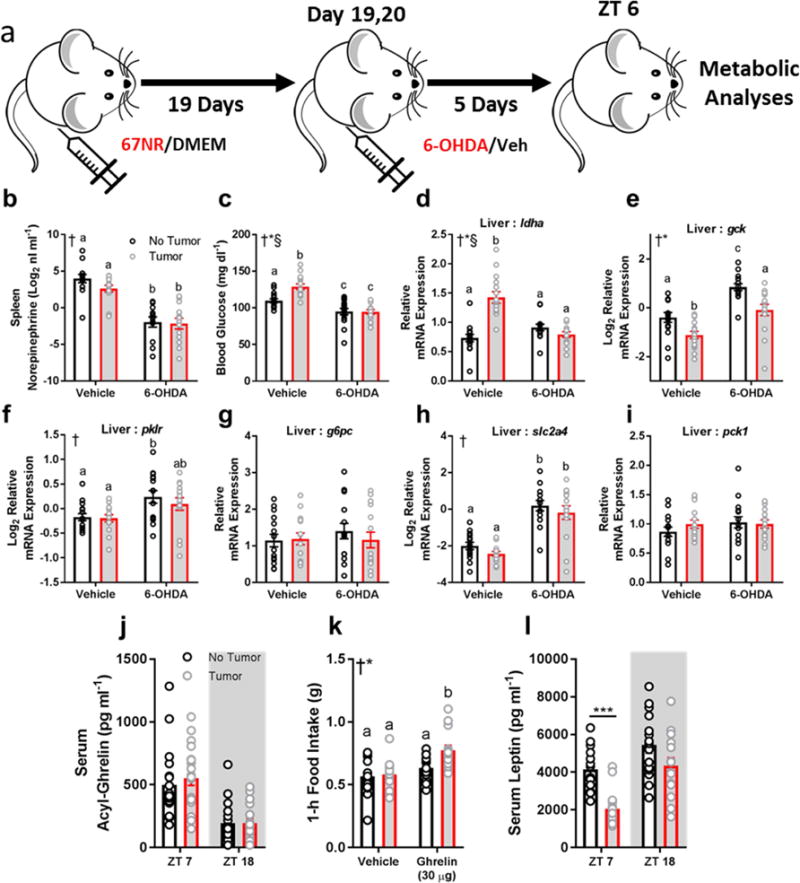
We investigated relationships among immune, metabolic, and sleep abnormalities in mice with non-metastatic mammary cancer. Tumor-bearing mice displayed interleukin-6 (IL-6)-mediated peripheral inflammation, coincident with altered hepatic glucose processing and sleep. Tumor-bearing mice were hyperphagic, had reduced serum leptin concentrations, and enhanced sensitivity to exogenous ghrelin. We tested whether these phenotypes were driven by inflammation using neutralizing monoclonal antibodies against IL-6; despite the reduction in IL-6 signaling, metabolic and sleep abnormalities persisted. We next investigated neural populations coupling metabolism and sleep, and observed altered activity within lateral-hypothalamic hypocretin/orexin (HO) neurons. We used a dual HO-receptor antagonist to test whether increased HO signaling was causing metabolic abnormalities. This approach rescued metabolic abnormalities and enhanced sleep quality in tumor-bearing mice. Peripheral sympathetic denervation prevented tumor-induced increases in serum glucose. Our results link metabolic and sleep abnormalities via the HO system, and provide evidence that central neuromodulators contribute to tumor-induced changes in metabolism.
Keywords: IL-6; breast cancer; ghrelin; glucose; hypocretin/orexin; leptin; sleep.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Cancer linked to sleep and metabolic disruption.Nat Rev Endocrinol. 2018 Aug;14(8):440. doi: 10.1038/s41574-018-0043-z. Nat Rev Endocrinol. 2018. PMID: 29907824 No abstract available.
-
Tumours trigger systemic disruption.Nat Rev Cancer. 2018 Aug;18(8):468-469. doi: 10.1038/s41568-018-0040-5. Nat Rev Cancer. 2018. PMID: 29930255 No abstract available.
-
Cancer: The Tumor-Driven Disease of the Host.Cell Metab. 2018 Jul 3;28(1):5-6. doi: 10.1016/j.cmet.2018.06.016. Cell Metab. 2018. PMID: 29972797
References
-
- Aslakson CJ, Miller FR. Selective Events in the Metastatic Process Defined by Analysis of the Sequential Dissemination of Subpopulations of a Mouse Mammary Tumor. Cancer Res. 1992;52:1399–1405. - PubMed
-
- Bedrosian TA, Fonken LK, Walton JC, Haim A, Nelson RJ. Dim light at night provokes depression-like behaviors and reduces CA1 dendritic spine density in female hamsters. Psychoneuroendocrinology. 2011;36:1062–1069. - PubMed
-
- Ben-Shaanan TL, Azulay-Debby H, Dubovik T, Starosvetsky E, Korin B, Schiller M, Green NL, Admon Y, Hakim F, Shen-Orr SS, et al. Activation of the reward system boosts innate and adaptive immunity. Nat Med. 2016;22:940–944. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
