

Breast cancer stem cell-like cells generated during TGFβ-induced EMT are radioresistant
- PMID: 29805752
- PMCID: PMC5955125
- DOI: 10.18632/oncotarget.25240
Breast cancer stem cell-like cells generated during TGFβ-induced EMT are radioresistant
Abstract
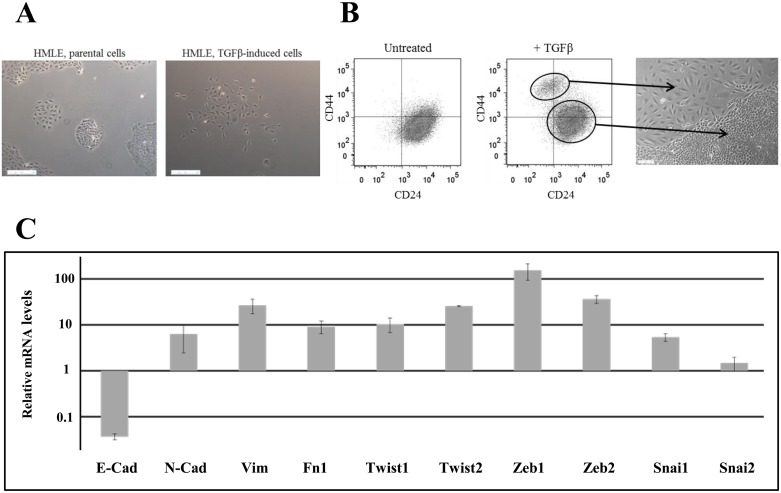
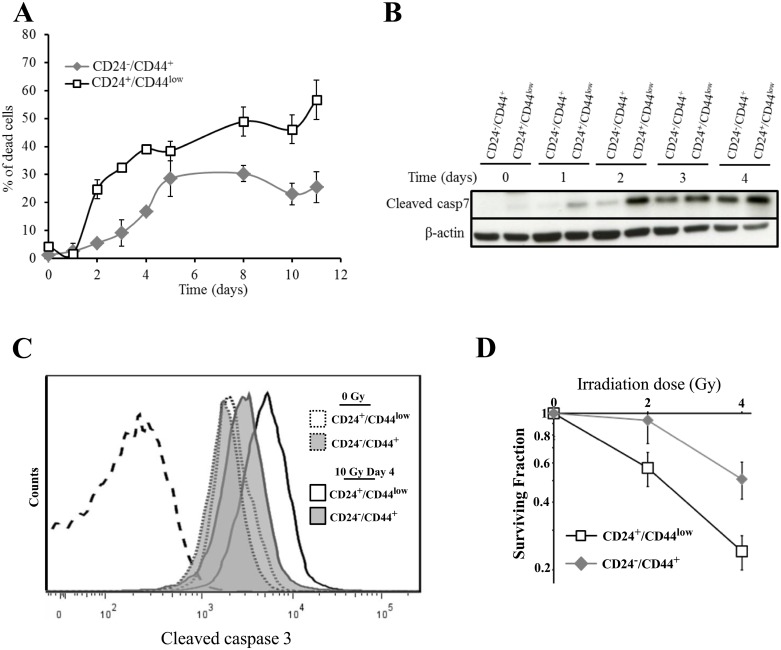
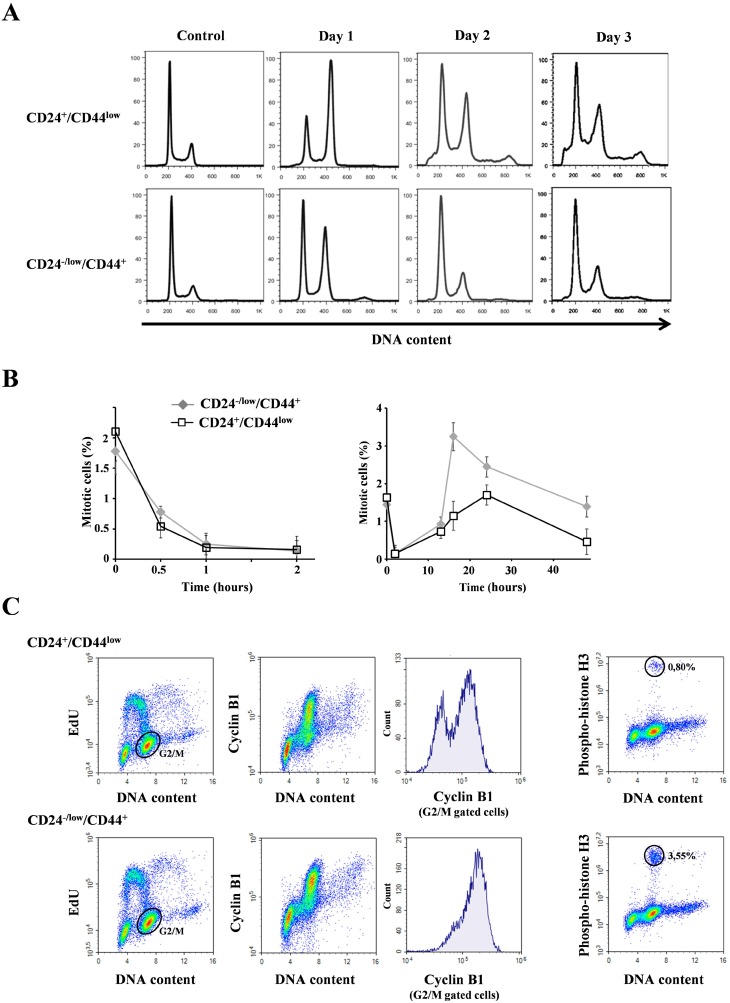
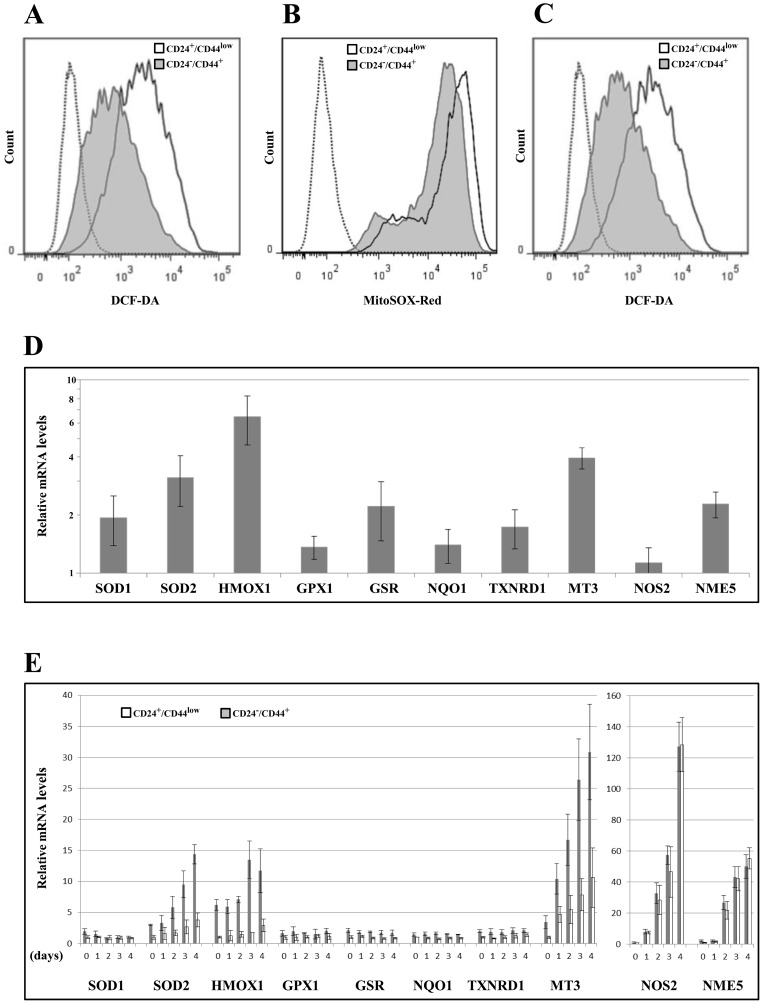
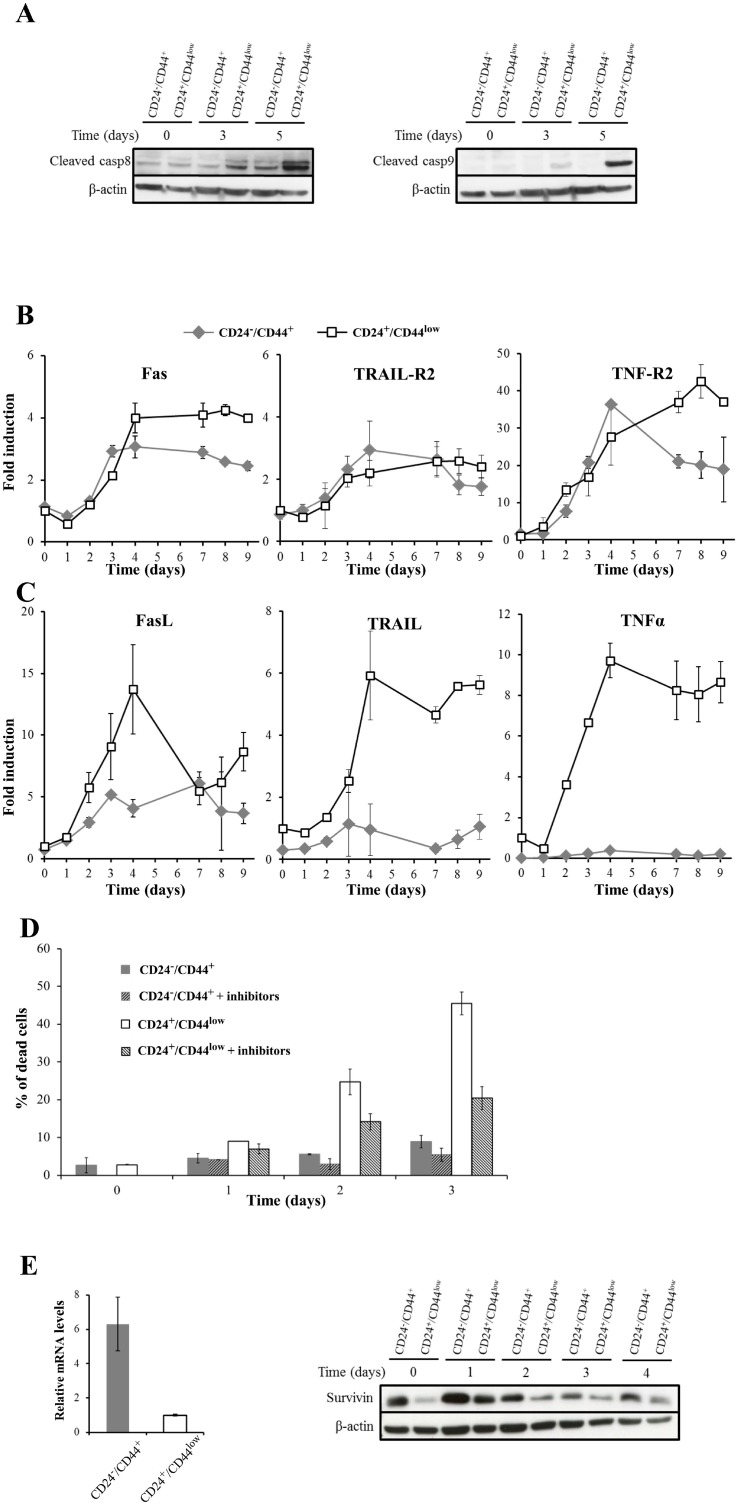
Failure of conventional antitumor therapy is commonly associated with cancer stem cells (CSCs), which are often defined as inherently resistant to radiation and chemotherapeutic agents. However, controversy about the mechanisms involved in the radiation response remains and the inherent intrinsic radioresistance of CSCs has also been questioned. These discrepancies observed in the literature are strongly associated with the cell models used. In order to clarify these contradictory observations, we studied the radiosensitivity of breast CSCs using purified CD24-/low/CD44+ CSCs and their corresponding CD24+/CD44low non-stem cells. These cells were generated after induction of the epithelial-mesenchymal transition (EMT) by transforming growth factor β (TGFβ) in immortalized human mammary epithelial cells (HMLE). Consequently, these 2 cellular subpopulations have an identical genetic background, their differences being related exclusively to TGFβ-induced cell reprogramming. We showed that mesenchymal CD24-/low/CD44+ CSCs are more resistant to radiation compared with CD24+/CD44low parental cells. Cell cycle distribution and free radical scavengers, but not DNA repair efficiency, appeared to be intrinsic determinants of cellular radiosensitivity. Finally, for the first time, we showed that reduced radiation-induced activation of the death receptor pathways (FasL, TRAIL and TNF-α) at the transcriptional level was a key causal event in the radioresistance of CD24-/low/CD44+ cells acquired during EMT.
Keywords: breast cancer; cancer stem cells; epithelial-mesenchymal transition; radioresistance.
Conflict of interest statement
CONFLICTS OF INTEREST None declared.
Figures

References
-
- Gupta PB, Chaffer CL, Weinberg RA. Cancer stem cells: mirage or reality? Nat Med. 2009;15:1010–1012. - PubMed
-
- Bao S, Wu Q, McLendon RE, Hao Y, Shi Q, Hjelmeland AB, Dewhirst MW, Bigner DD, Rich JN. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature. 2006;444:756–760. - PubMed
-
- Li X, Lewis MT, Huang J, Gutierrez C, Osborne CK, Wu MF, Hilsenbeck SG, Pavlick A, Zhang X, Chamness GC, Wong H, Rosen J, Chang JC. Intrinsic resistance of tumorigenic breast cancer cells to chemotherapy. J Natl Cancer Inst. 2008;100:672–679. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
