

Hematopoietic stem cell fate through metabolic control
- PMID: 29807063
- PMCID: PMC6084457
- DOI: 10.1016/j.exphem.2018.05.005
Hematopoietic stem cell fate through metabolic control
Abstract
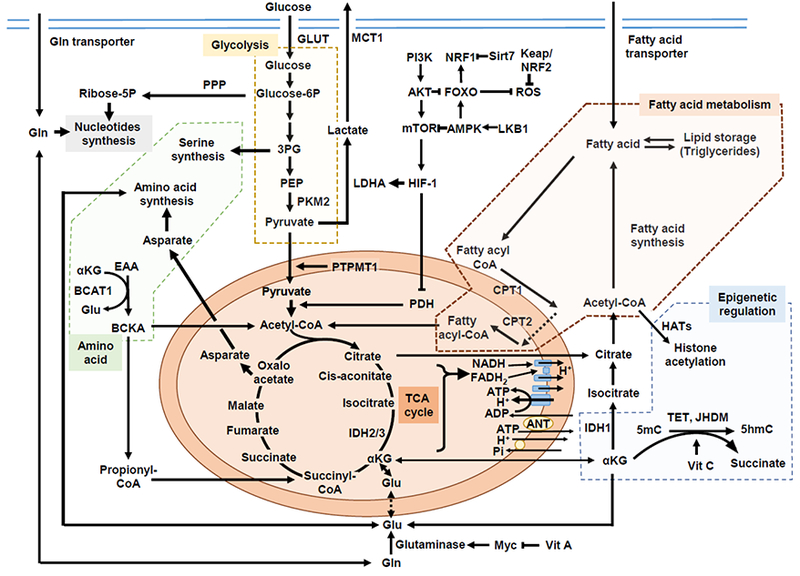
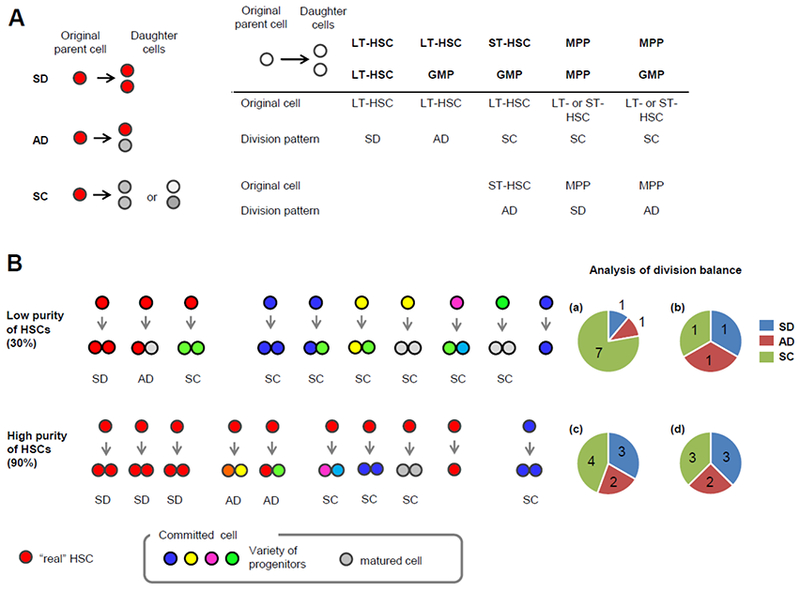
Hematopoietic stem cells maintain a quiescent state in the bone marrow to preserve their self-renewal capacity, but also undergo cell divisions as required. Organelles such as the mitochondria sustain cumulative damage during these cell divisions and this damage may eventually compromise the cells' self-renewal capacity. Hematopoietic stem cell divisions result in either self-renewal or differentiation, with the balance between the two affecting hematopoietic homeostasis directly; however, the heterogeneity of available hematopoietic stem cell-enriched fractions, together with the technical challenges of observing hematopoietic stem cell behavior, has long hindered the analysis of individual hematopoietic stem cells and prevented the elucidation of this process. Recent advances in genetic models, metabolomics analyses, and single-cell approaches have revealed the contributions made to hematopoietic stem cell self-renewal by metabolic cues, mitochondrial biogenesis, and autophagy/mitophagy, which have highlighted mitochondrial quality control as a key factor in the equilibrium of hematopoietic stem cells. A deeper understanding of precisely how specific modes of metabolism control hematopoietic stem cells fate at the single-cell level is therefore not only of great biological interest, but will also have clear clinical implications for the development of therapies for hematological diseases.
Copyright © 2018 ISEH -- Society for Hematology and Stem Cells. Published by Elsevier Inc. All rights reserved.
Figures

References
-
- McCulloch EA & Till JE The radiation sensitivity of normal mouse bone marrow cells, determined by quantitative marrow transplantation into irradiated mice. Radiat Res 13, 115–125 (1960). - PubMed
-
- Morrison SJ, Shah NM & Anderson DJ Regulatory mechanisms in stem cell biology. Cell 88, 287–298 (1997). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
