

Review
doi: 10.1038/s41594-018-0075-z.
Epub 2018 May 28.
RPA and RAD51: fork reversal, fork protection, and genome stability
Affiliations
- PMID: 29807999
- PMCID: PMC6006513
- DOI: 10.1038/s41594-018-0075-z
Item in Clipboard
Review
RPA and RAD51: fork reversal, fork protection, and genome stability
Nat Struct Mol Biol.
2018 Jun.
Abstract
Replication protein A (RPA) and RAD51 are DNA-binding proteins that help maintain genome stability during DNA replication. These proteins regulate nucleases, helicases, DNA translocases, and signaling proteins to control replication, repair, recombination, and the DNA damage response. Their different DNA-binding mechanisms, enzymatic activities, and binding partners provide unique functionalities that cooperate to ensure that the appropriate activities are deployed at the right time to overcome replication challenges. Here we review and discuss the latest discoveries of the mechanisms by which these proteins work to preserve genome stability, with a focus on their actions in fork reversal and fork protection.
Figures
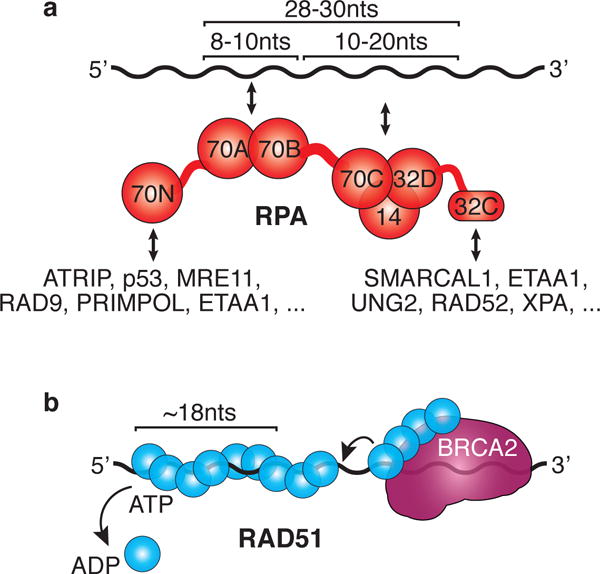
(A) RPA uses modular domain architecture to facilitate dynamic DNA and protein interactions. The size of the ssDNA that is bound by the RPA DNA binding domains is indicated as well as the major protein-protein interaction domains with example binding partners. (B) RAD51 forms a protein-ssDNA filament with the help of BRCA2 and is regulated by its ATPase activity. BRCA2 uses its own ssDNA binding activity to help deliver RAD51 and displace RPA (arrow). ATP hydrolysis promotes filament disassembly.
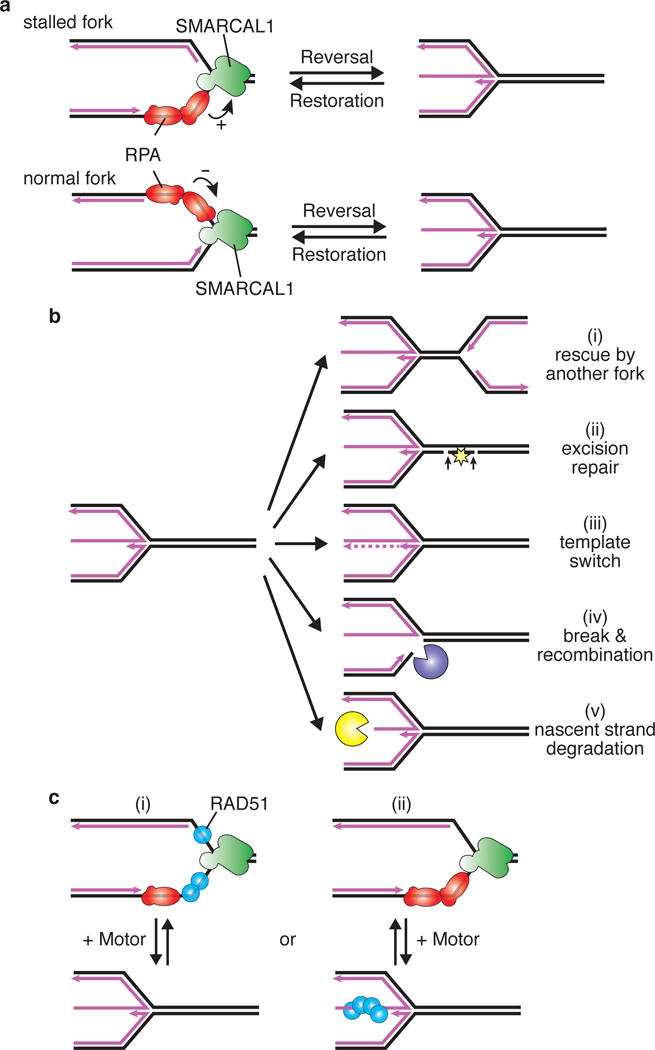
(A) RPA stimulates SMARCAL1 to catalyze fork reversal when bound to the leading strand template “stalled fork” but inhibits SMARCAL1 when bound to the lagging strand “normal fork”. The polarity difference of RPA bound to these substrates is illustrated since that contributes to SMARCAL1 regulation. Pink lines indicate nascent DNA while template DNA is illustrated in black. (B) Reversed forks are intermediates in fork stabilization and restart mechanisms, but are also susceptible to nuclease processing. Five outcomes of fork reversal are illustrated and described further in the text. (C) Two models for how RAD51 may promote fork reversal. First, RAD51 may bind the template ssDNA of the stalled fork to cooperate with motor proteins like SMARCAL1 and ZRANB3 to drive reversal. Second, RAD51 could capture the reversed fork, driving an equilibrium towards fork reversal.
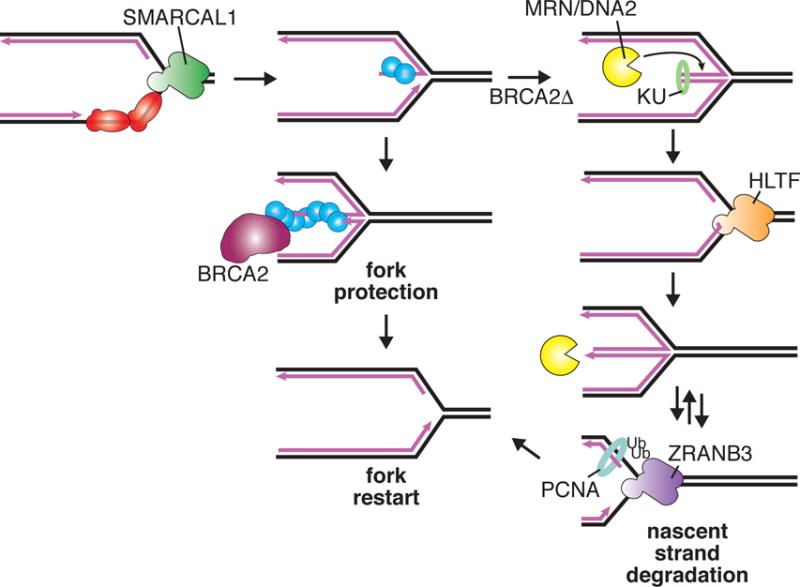
When RAD51-DNA filaments are not stabilized, nucleases including MRN and DNA2 can access the end of the reversed arm. Limited nascent strand degradation could be beneficial to remove end-binding proteins and allow fork restart. However, in the context of a persistent fork block, multiple rounds of end processing and reversal by motor proteins with different substrate specificities yields extensive degradation and repeated opportunities for mistakes that generate chromosomal abnormalities. Depicted is a speculative model of the motor proteins acting sequentially. Ub = ubiquitin.
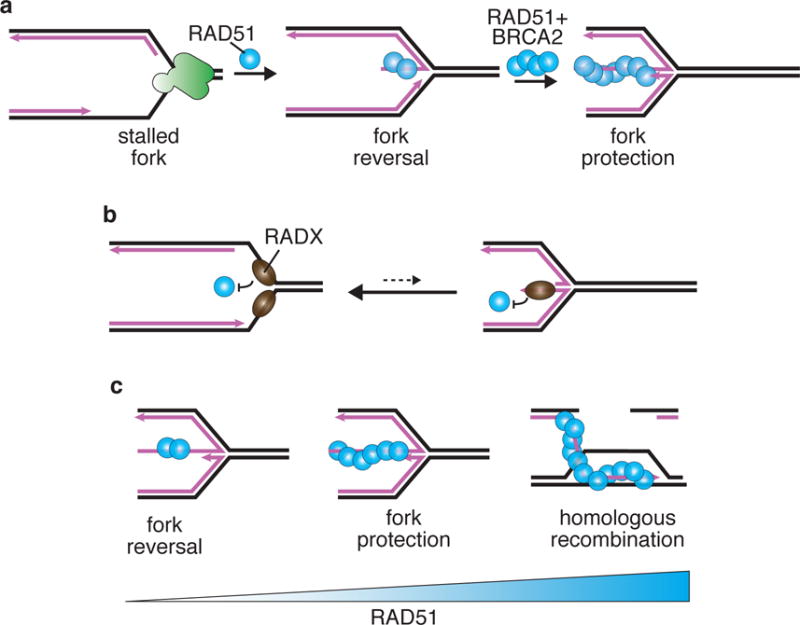
(A) RAD51 is needed for both fork reversal and fork protection. Fork reversal does not require BRCA2 and may require less cellular RAD51 protein than fork protection. (B) During normal replication, transient exposures of ssDNA at an elongating replication fork may be shielded from RAD51 by RADX to prevent unnecessary fork reversal. (B) Increasing amounts of RAD51 protein/activity may be needed for fork reversal, fork protection, and HR.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
