

Prohibitin is a positive modulator of mitochondrial function in PC12 cells under oxidative stress
- PMID: 29808474
- PMCID: PMC6105506
- DOI: 10.1111/jnc.14472
Prohibitin is a positive modulator of mitochondrial function in PC12 cells under oxidative stress
Abstract
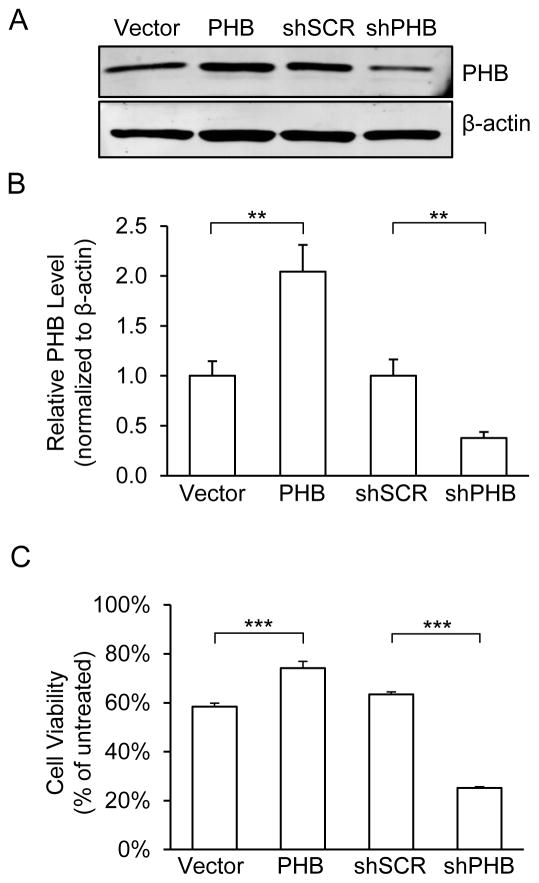
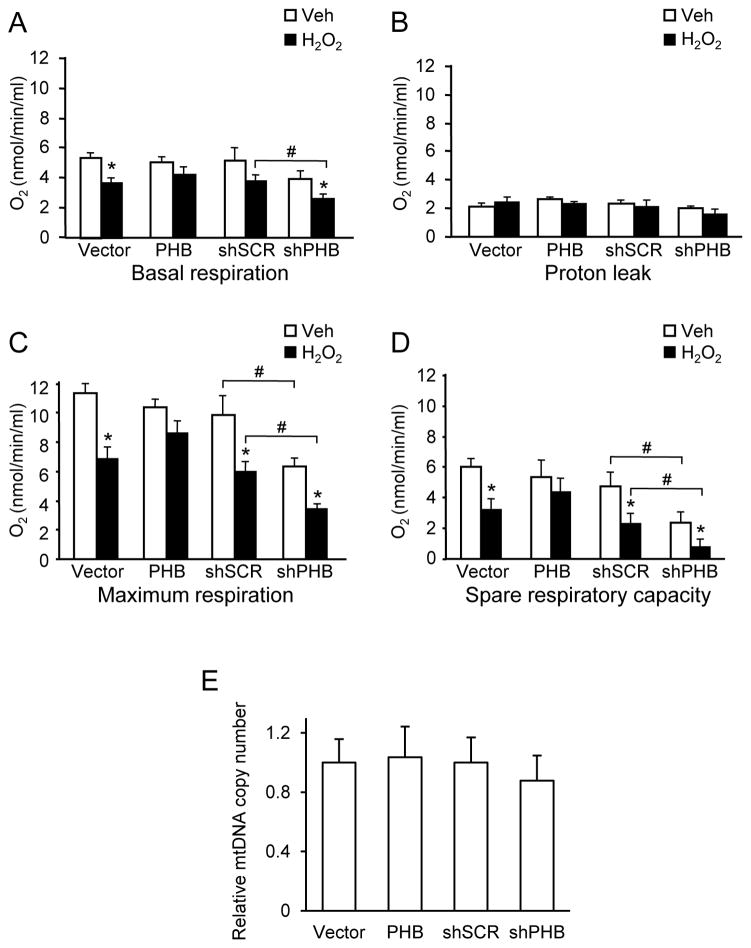
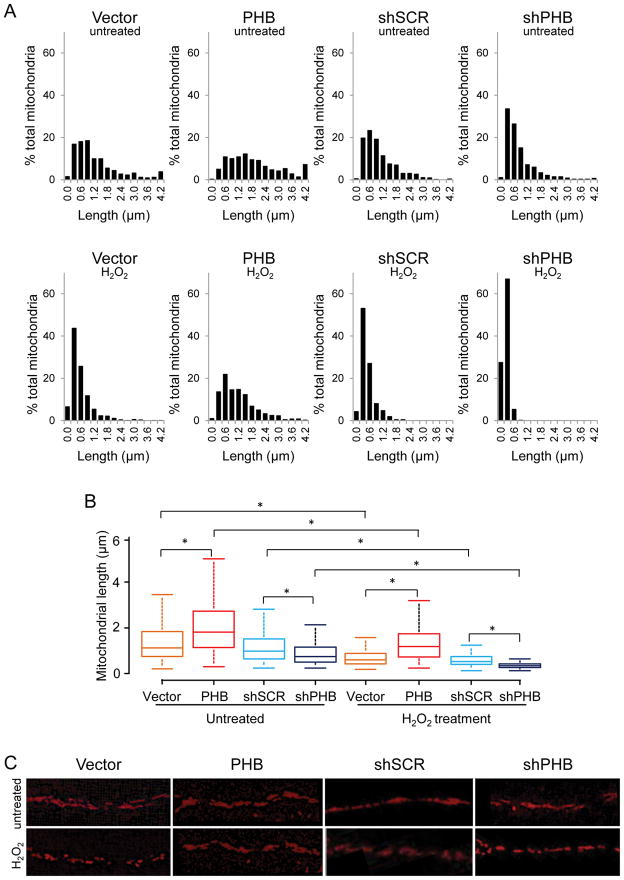
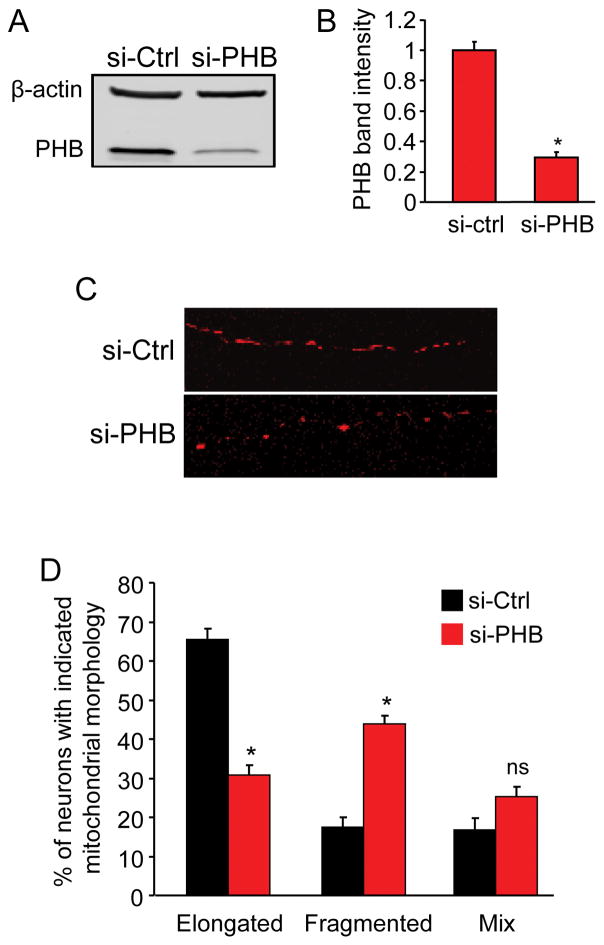
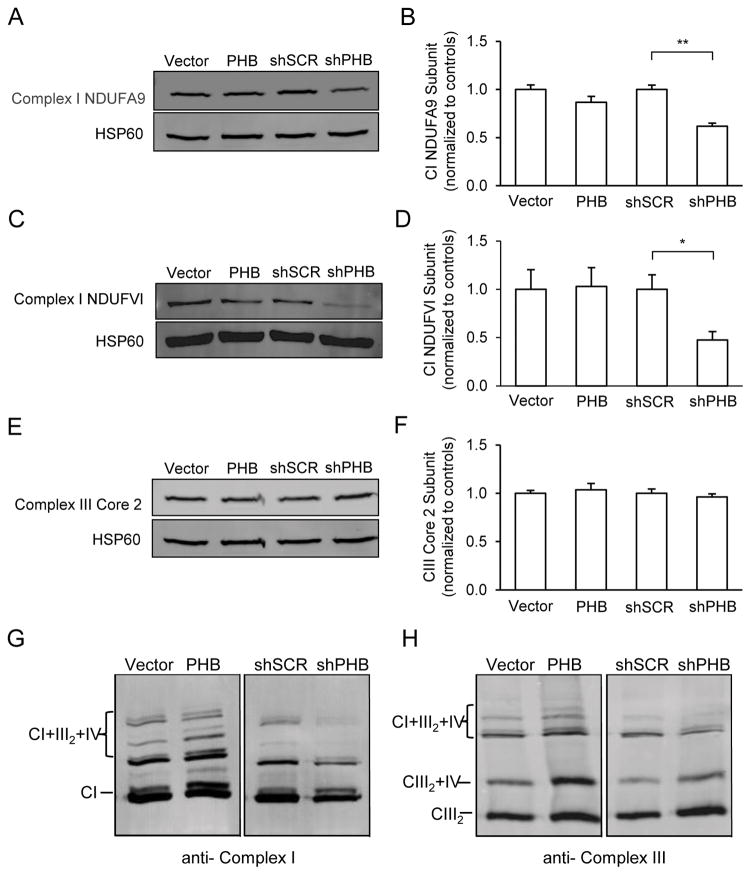
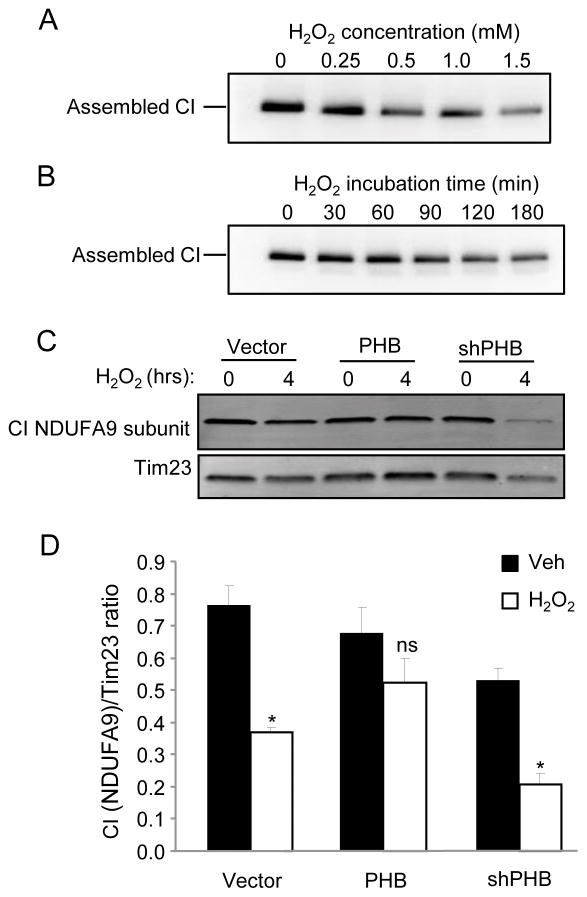
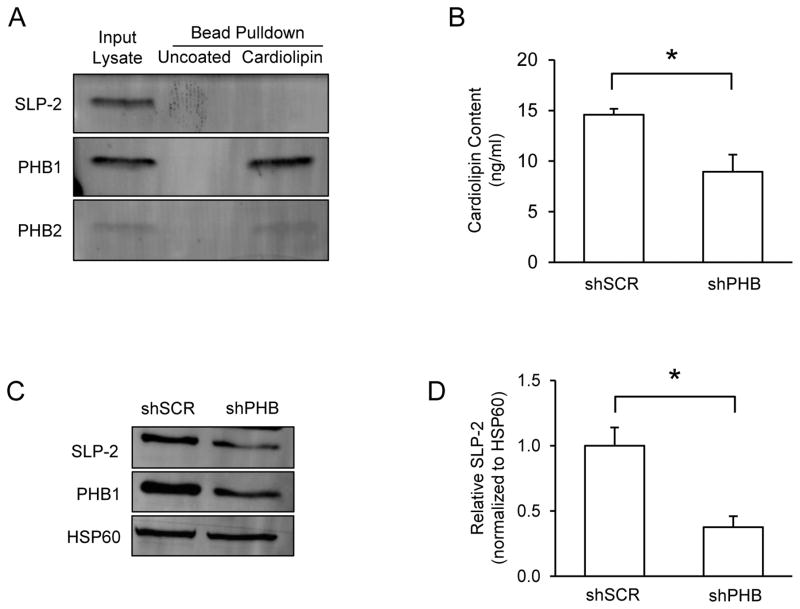
Prohibitin (PHB) is a ubiquitously expressed and evolutionarily conserved mitochondrial protein with multiple functions. We have recently shown that PHB up-regulation offers robust protection against neuronal injury in models of cerebral ischemia in vitro and in vivo, but the mechanism by which PHB affords neuroprotection remains to be elucidated. Here, we manipulated PHB expression in PC12 neural cells to investigate its impact on mitochondrial function and the mechanisms whereby it protects cells exposed to oxidative stress. PHB over-expression promoted cell survival, whereas PHB down-regulation diminished cell viability. Functionally, manipulation of PHB levels did not affect basal mitochondrial respiration, but it increased spare respiratory capacity. Moreover, PHB over-expression preserved mitochondrial respiratory function of cells exposed to oxidative stress. Preserved respiratory capacity in differentiated PHB over-expressing cells exposed to oxidative stress was associated with an elongated mitochondrial morphology, whereas PHB down-regulation enhanced fragmentation. Mitochondrial complex I oxidative degradation was attenuated by PHB over-expression and increased in PHB knockdown cells. Changes in complex I degradation were associated with alterations of respiratory chain supercomplexes. Furthermore, we showed that PHB directly interacts with cardiolipin and that down-regulation of PHB results in loss of cardiolipin in mitochondria, which may contribute to destabilizing respiratory chain supercomplexes. Taken together, these data demonstrate that PHB modulates mitochondrial integrity and bioenergetics under oxidative stress, and suggest that the protective effect of PHB is mediated by stabilization of the mitochondrial respiratory machinery and its functional capacity, by the regulation of cardiolipin content. Open Data: Materials are available on https://cos.io/our-services/open-science-badges/ https://osf.io/93n6m/.
Keywords: PC12 cells; mitochondria; oxidative stress; prohibitin; respiratory chain complexes.
© 2018 International Society for Neurochemistry.
Conflict of interest statement
All authors claim no conflicts of interest.
Figures
References
-
- Acin-Perez R, Fernandez-Silva P, Peleato ML, Perez-Martos A, Enriquez JA. Respiratory active mitochondrial supercomplexes. Molecular cell. 2008;32:529–539. - PubMed
-
- Artal-Sanz M, Tsang WY, Willems EM, Grivell LA, Lemire BD, van der Spek H, Nijtmans LG. The mitochondrial prohibitin complex is essential for embryonic viability and germline function in Caenorhabditis elegans. The Journal of biological chemistry. 2003;278:32091–32099. - PubMed
-
- Ban T, Ishihara T, Kohno H, Saita S, Ichimura A, Maenaka K, Oka T, Mihara K, Ishihara N. Molecular basis of selective mitochondrial fusion by heterotypic action between OPA1 and cardiolipin. Nature cell biology. 2017;19:856–863. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
