

Zbtb7a is a transducer for the control of promoter accessibility by NF-kappa B and multiple other transcription factors
- PMID: 29813070
- PMCID: PMC5993293
- DOI: 10.1371/journal.pbio.2004526
Zbtb7a is a transducer for the control of promoter accessibility by NF-kappa B and multiple other transcription factors
Abstract
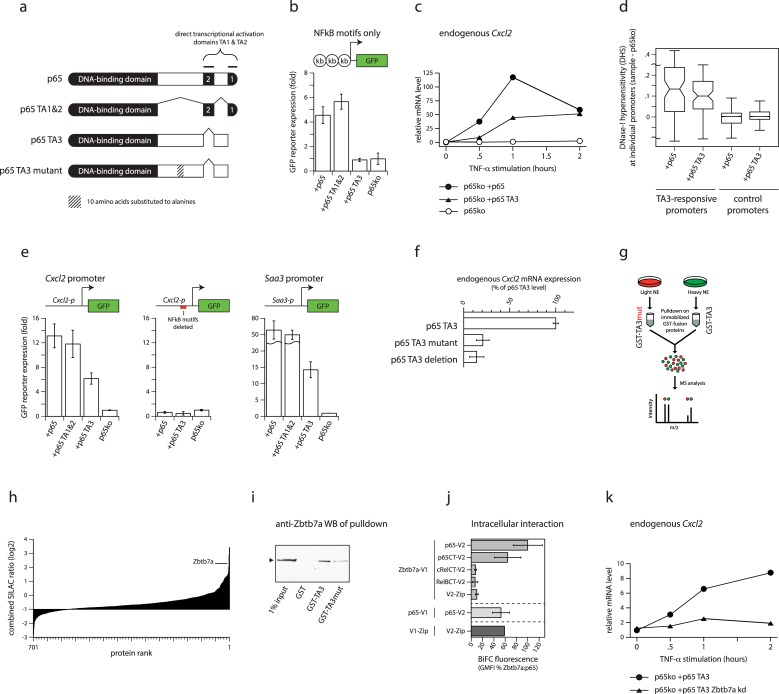
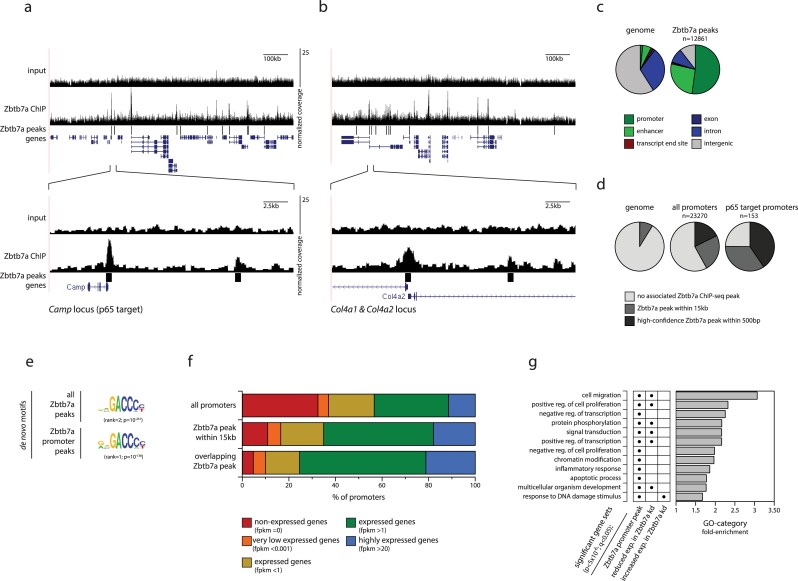
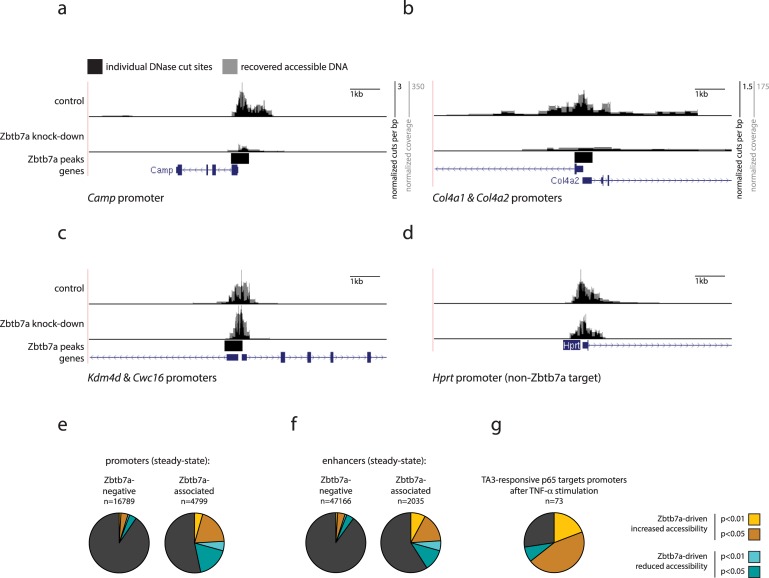
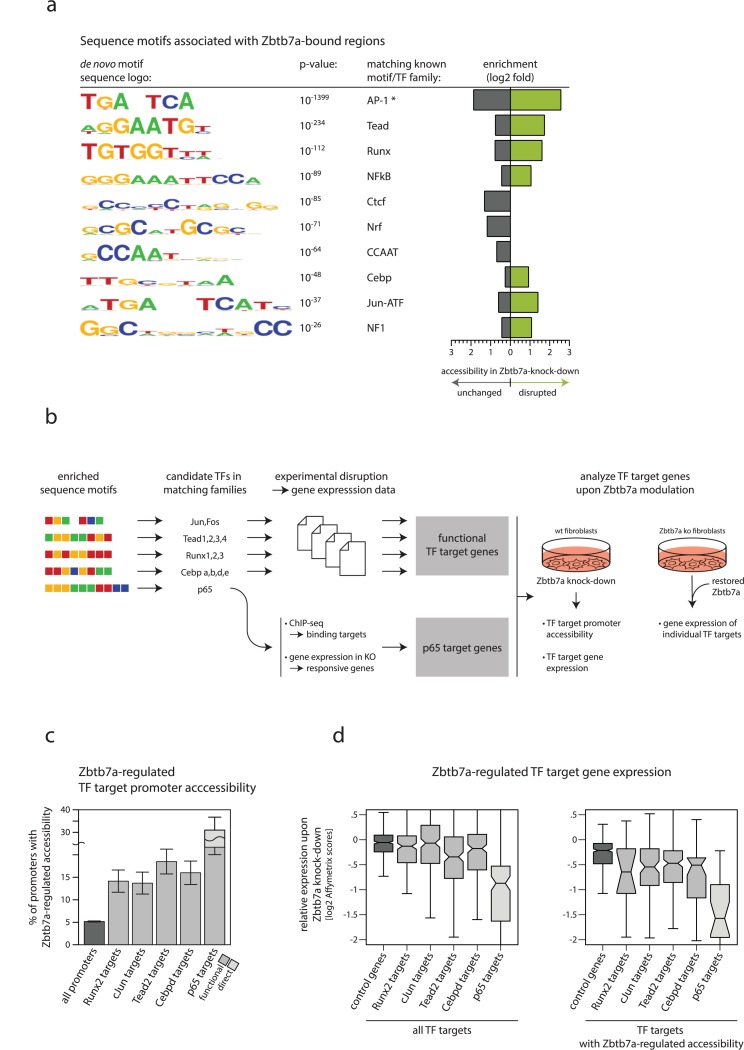
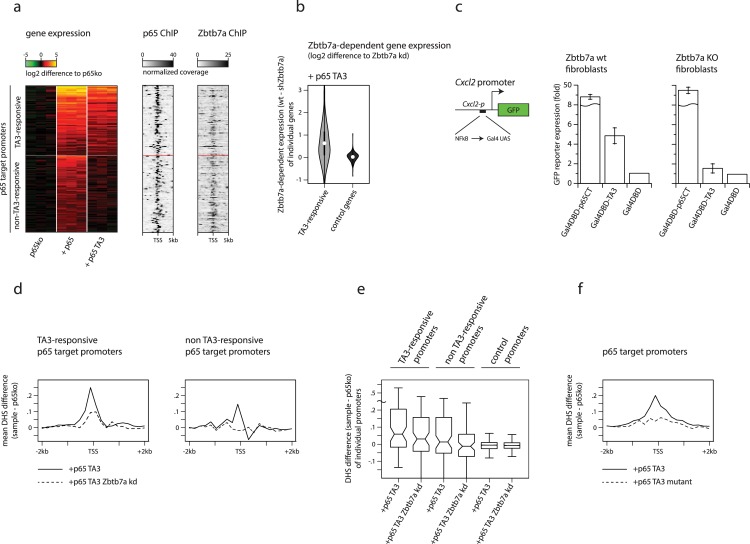
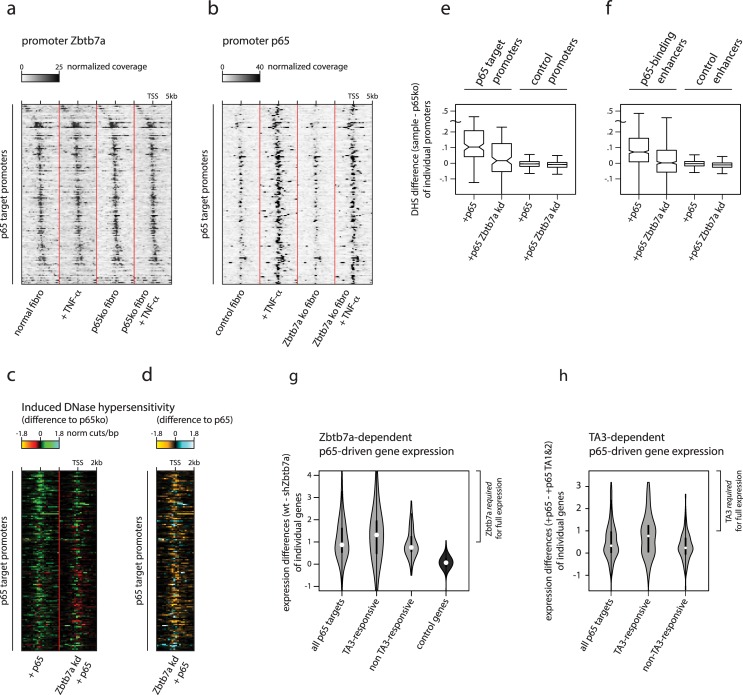
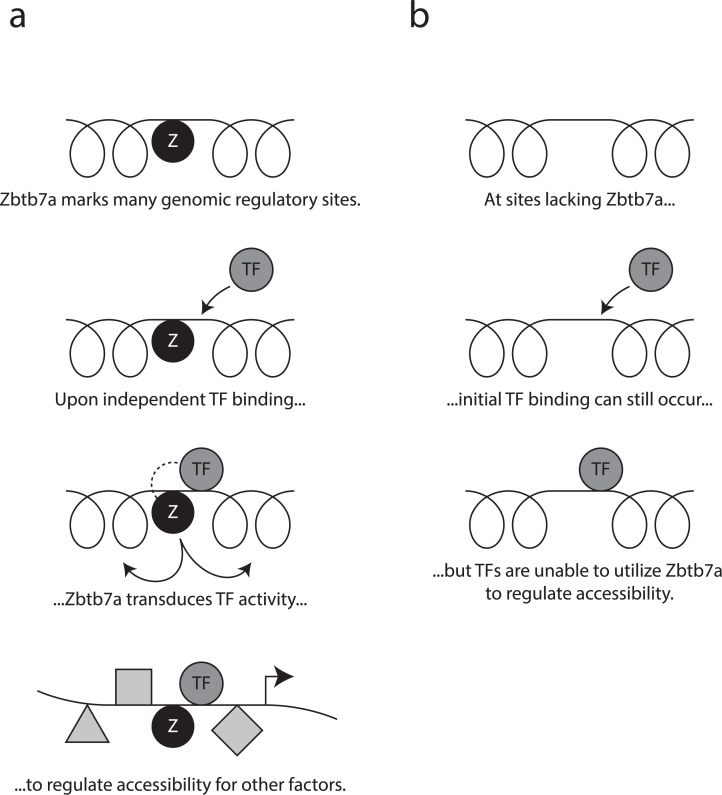
Gene expression in eukaryotes is controlled by DNA sequences at promoter and enhancer regions, whose accessibility for binding by regulatory proteins dictates their specific patterns of activity. Here, we identify the protein Zbtb7a as a factor required for inducible changes in accessibility driven by transcription factors (TFs). We show that Zbtb7a binds to a significant fraction of genomic promoters and enhancers, encompassing many target genes of nuclear factor kappa B (NFκB) p65 and a variety of other TFs. While Zbtb7a binding is not alone sufficient to directly activate promoters, it is required to enable TF-dependent control of accessibility and normal gene expression. Using p65 as a model TF, we show that Zbtb7a associates with promoters independently of client TF binding. Moreover, the presence of prebound Zbtb7a can specify promoters that are amenable to TF-induced changes in accessibility. Therefore, Zbtb7a represents a widely used promoter factor that transduces signals from other TFs to enable control of accessibility and regulation of gene expression.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Bell O, Tiwari VK, Thoma NH, Schubeler D (2011) Determinants and dynamics of genome accessibility. Nat Rev Genet 12: 554–564. doi: 10.1038/nrg3017 - DOI - PubMed
-
- Guertin MJ, Lis JT (2013) Mechanisms by which transcription factors gain access to target sequence elements in chromatin. Curr Opin Genet Dev 23: 116–123. doi: 10.1016/j.gde.2012.11.008 - DOI - PMC - PubMed
-
- Chen T, Dent SY (2014) Chromatin modifiers and remodellers: regulators of cellular differentiation. Nat Rev Genet 15: 93–106. doi: 10.1038/nrg3607 - DOI - PMC - PubMed
-
- Clapier CR, Cairns BR (2009) The biology of chromatin remodeling complexes. Annu Rev Biochem 78: 273–304. doi: 10.1146/annurev.biochem.77.062706.153223 - DOI - PubMed
-
- Buenrostro JD, Giresi PG, Zaba LC, Chang HY, Greenleaf WJ (2013) Transposition of native chromatin for fast and sensitive epigenomic profiling of open chromatin, DNA-binding proteins and nucleosome position. Nat Methods 10: 1213–1218. doi: 10.1038/nmeth.2688 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
