

Polymorphic factor H-binding activity of CspA protects Lyme borreliae from the host complement in feeding ticks to facilitate tick-to-host transmission
- PMID: 29813137
- PMCID: PMC5993331
- DOI: 10.1371/journal.ppat.1007106
Polymorphic factor H-binding activity of CspA protects Lyme borreliae from the host complement in feeding ticks to facilitate tick-to-host transmission
Abstract
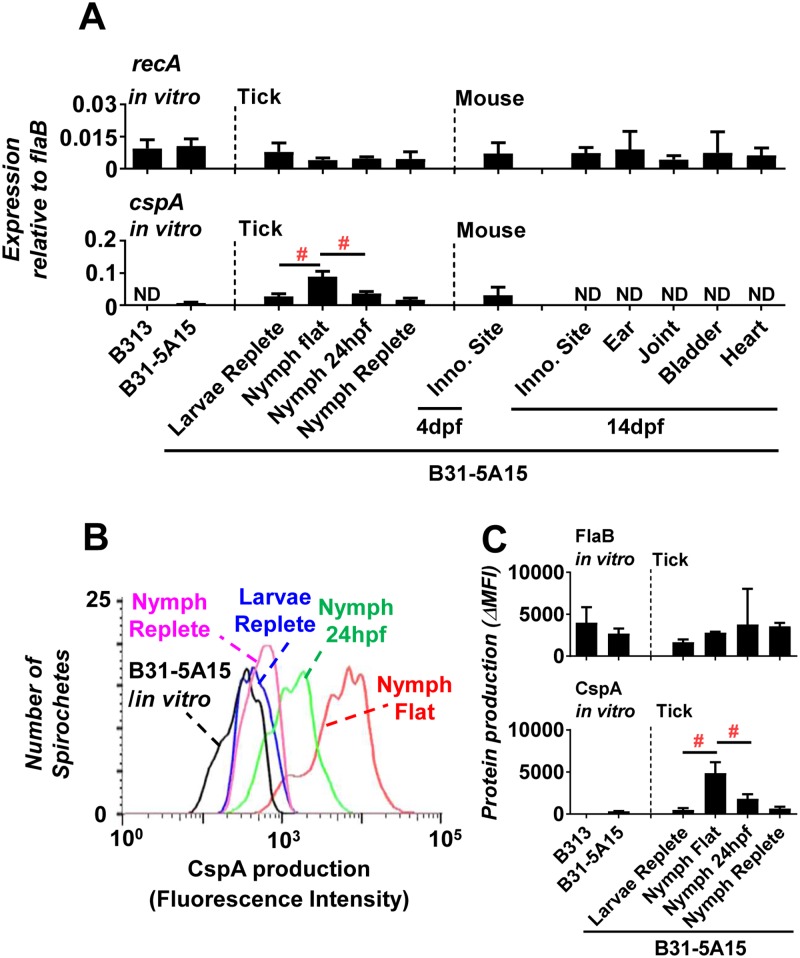
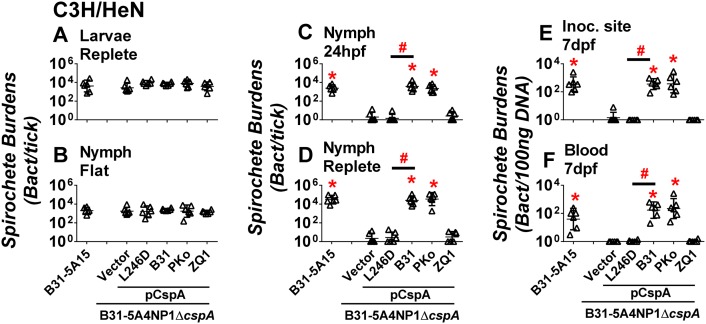
Borrelia burgdorferi sensu lato (Bbsl), the causative agent of Lyme disease, establishes an initial infection in the host's skin following a tick bite, and then disseminates to distant organs, leading to multisystem manifestations. Tick-to-vertebrate host transmission requires that Bbsl survives during blood feeding. Complement is an important innate host defense in blood and interstitial fluid. Bbsl produces a polymorphic surface protein, CspA, that binds to a complement regulator, Factor H (FH) to block complement activation in vitro. However, the role that CspA plays in the Bbsl enzootic cycle remains unclear. In this study, we demonstrated that different CspA variants promote spirochete binding to FH to inactivate complement and promote serum resistance in a host-specific manner. Utilizing a tick-to-mouse transmission model, we observed that a cspA-knockout B. burgdorferi is eliminated from nymphal ticks in the first 24 hours of feeding and is unable to be transmitted to naïve mice. Conversely, ectopically producing CspA derived from B. burgdorferi or B. afzelii, but not B. garinii in a cspA-knockout strain restored spirochete survival in fed nymphs and tick-to-mouse transmission. Furthermore, a CspA point mutant, CspA-L246D that was defective in FH-binding, failed to survive in fed nymphs and at the inoculation site or bloodstream in mice. We also allowed those spirochete-infected nymphs to feed on C3-/- mice that lacked functional complement. The cspA-knockout B. burgdorferi or this mutant strain complemented with cspA variants or cspA-L246D was found at similar levels as wild type B. burgdorferi in the fed nymphs and mouse tissues. These novel findings suggest that the FH-binding activity of CspA protects spirochetes from complement-mediated killing in fed nymphal ticks, which ultimately allows Bbsl transmission to mammalian hosts.
Conflict of interest statement
The authors have declared no competing interests exist.
Figures
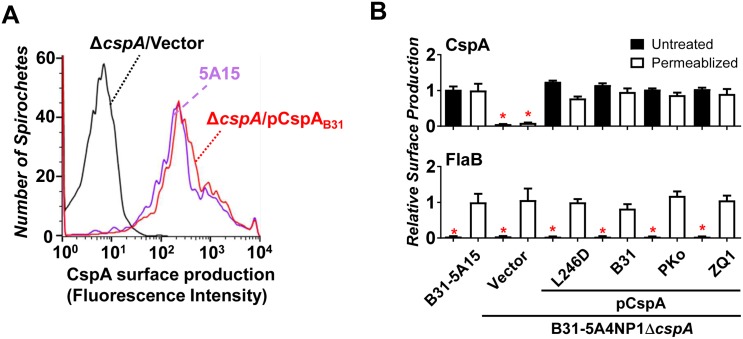
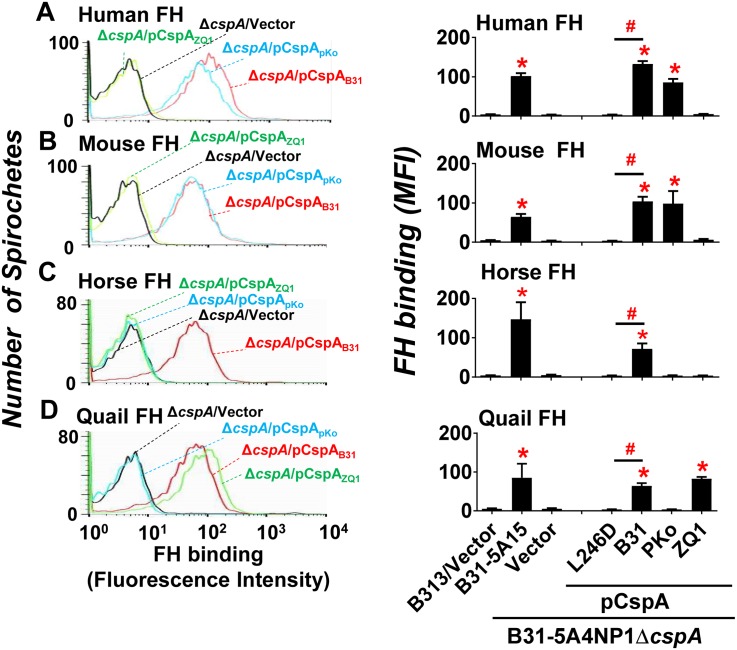
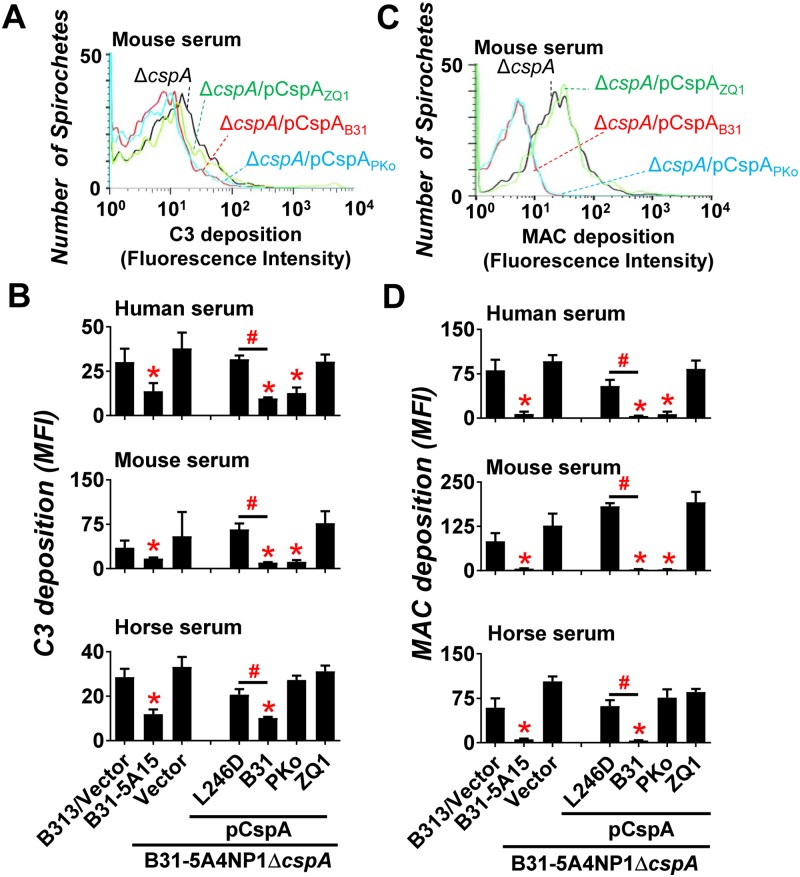
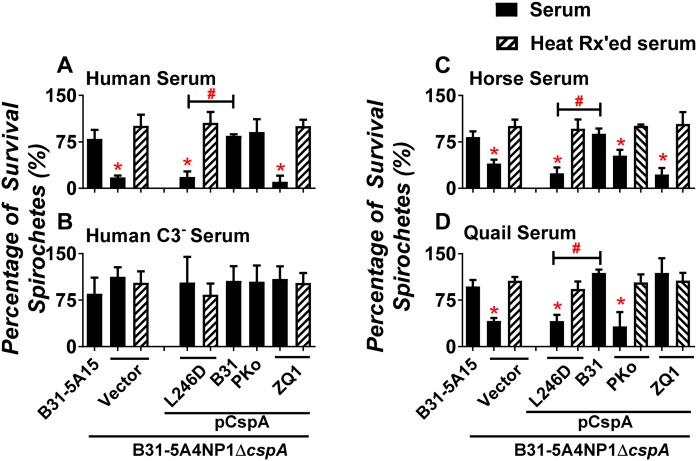


References
-
- Steere AC, Strle F, Wormser GP, Hu LT, Branda JA, Hovius JW, et al. Lyme borreliosis. Nat Rev Dis Primers. 2016;2:16090 doi: 10.1038/nrdp.2016.90 . - DOI - PMC - PubMed
-
- Radolf JD, Caimano MJ, Stevenson B, Hu LT. Of ticks, mice and men: understanding the dual-host lifestyle of Lyme disease spirochaetes. Nat Rev Microbiol. 2012;10(2):87–99. doi: 10.1038/nrmicro2714 . - DOI - PMC - PubMed
-
- Rosa PA, Tilly K, Stewart PE. The burgeoning molecular genetics of the Lyme disease spirochaete. Nat Rev Microbiol. 2005;3(2):129–43. doi: 10.1038/nrmicro1086 . - DOI - PubMed
-
- Steere AC, Coburn J, Glickstein L. The emergence of Lyme disease. J Clin Invest. 2004;113(8):1093–101. doi: 10.1172/JCI21681 . - DOI - PMC - PubMed
-
- Brisson D, Drecktrah D, Eggers CH, Samuels DS. Genetics of Borrelia burgdorferi. Annu Rev Genet. 2012;46:515–36. doi: 10.1146/annurev-genet-011112-112140 . - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
